
Статьи. Предметно
Аксон Ходхкина-Хаксли-Катца с сальтаторным механихмом
Модель базируется на электрохимическом уравнении (3). Мембранный потенциал будем отсчитывать от его уровня ![]() в состоянии покоя, т.е. положим
в состоянии покоя, т.е. положим ![]() , или же
, или же ![]() . Аналогично введем обозначения для отсчитываемых от
. Аналогично введем обозначения для отсчитываемых от ![]() равновесных электрохимических потенциалов
равновесных электрохимических потенциалов ![]() и
и ![]() . Ясно, что сохраняются разности:
. Ясно, что сохраняются разности: ![]() и
и ![]() . В создании мембранного потенциала кроме ионов
. В создании мембранного потенциала кроме ионов ![]() и
и ![]() принимают участие и другие ионы, в частности, ионы
принимают участие и другие ионы, в частности, ионы ![]() . Обозначим их суммарный электрохимический потенциал через
. Обозначим их суммарный электрохимический потенциал через ![]() , соответственно
, соответственно ![]() . В уравнение (3) для баланса мембранных токов следует добавить слагаемое
. В уравнение (3) для баланса мембранных токов следует добавить слагаемое ![]() . Данный ток называется током утечки. Для гигантского аксона кальмара А. Ходжкиным и А. Хаксли были экспериментально найдены значения:
. Данный ток называется током утечки. Для гигантского аксона кальмара А. Ходжкиным и А. Хаксли были экспериментально найдены значения: ![]() мв,
мв, ![]() мвт,
мвт, ![]() мвт.
мвт.
Электрохимическое уравнение (3) суть обыкновенное дифференциальное уравнение первого порядка. Как показано выше, его коэффициенты следует считать функциями от мембранного потенциала ![]() . По биологическому смыслу решения уравнения ограничены на положительной полуоси
. По биологическому смыслу решения уравнения ограничены на положительной полуоси ![]() . Можно доказать, что все решения такого уравнения монотонно стремятся к состояниям равновесия. Объяснить в рамках одного электрохимического уравнения генерацию потенциала действия невозможно.
. Можно доказать, что все решения такого уравнения монотонно стремятся к состояниям равновесия. Объяснить в рамках одного электрохимического уравнения генерацию потенциала действия невозможно.
Из приведенных выше построений следует, проводимости ![]() и
и ![]() следует считать функциями не только мембранного потенциала
следует считать функциями не только мембранного потенциала ![]() , но и времени. Нижеследующие построения А. Ходжкина и А. Хаксли во многом носят феноменологический характер, т.е. не имееют под собой прочной теоретической основы и опираются на ряд допущений и догадок. Рассмотрим натриевую проводимость. Для описания ее зависимости от времени и мембранного потенциала А. Ходжкин и А. Хаксли ввели две новые переменные
, но и времени. Нижеследующие построения А. Ходжкина и А. Хаксли во многом носят феноменологический характер, т.е. не имееют под собой прочной теоретической основы и опираются на ряд допущений и догадок. Рассмотрим натриевую проводимость. Для описания ее зависимости от времени и мембранного потенциала А. Ходжкин и А. Хаксли ввели две новые переменные ![]() и
и ![]() , которые характеризуют процессы активации и инактивации натриевых каналов. В интерпретации авторов
, которые характеризуют процессы активации и инактивации натриевых каналов. В интерпретации авторов ![]() -вероятность открытия
-вероятность открытия ![]() -ворот,
-ворот, ![]() -вероятность закрытия
-вероятность закрытия ![]() -ворот, или же -отношение числа открытых и закрытых ворот соответствующего типа к числу каналов на поверхности мембраны единичной площади. Авторы модели положили:
-ворот, или же -отношение числа открытых и закрытых ворот соответствующего типа к числу каналов на поверхности мембраны единичной площади. Авторы модели положили: ![]() , где коэффициент
, где коэффициент ![]() был назван максимальной натриевой проводимостью. Предложенное А. Ходжкиным и А. Хаксли обоснование вида зависимости в настоящее время выглядит искуственным. Методом подбора были выписаны линейные обыкновенные уравнения первого порядка для переменных
был назван максимальной натриевой проводимостью. Предложенное А. Ходжкиным и А. Хаксли обоснование вида зависимости в настоящее время выглядит искуственным. Методом подбора были выписаны линейные обыкновенные уравнения первого порядка для переменных ![]() и
и ![]() . При этом в уравнение для
. При этом в уравнение для ![]() не входит переменная
не входит переменная ![]() , а в уравнение для
, а в уравнение для ![]() не входит
не входит ![]() . Коэффициенты уравнений зависят только от мембранного потенциала
. Коэффициенты уравнений зависят только от мембранного потенциала ![]() . Более менее убедительное обоснование необходимости именно такой зависимости отсутствует.
. Более менее убедительное обоснование необходимости именно такой зависимости отсутствует.
Для описания изменения калиевой проводимости ![]() А. Ходжкиным и А. Хаксли была введена функция
А. Ходжкиным и А. Хаксли была введена функция ![]() . Она интерпретировалась как вероятность открытия
. Она интерпретировалась как вероятность открытия ![]() -ворот для каливых каналов, т.е. как вероятность активации последних. Напомним, что по гипотезе А. Ходжкина и А. Хаксли инактивационные ворота для каливых каналов отсутствуют. Авторы опытным путем подобрали линейное обыкновенное дифференциальное уравнение для
-ворот для каливых каналов, т.е. как вероятность активации последних. Напомним, что по гипотезе А. Ходжкина и А. Хаксли инактивационные ворота для каливых каналов отсутствуют. Авторы опытным путем подобрали линейное обыкновенное дифференциальное уравнение для ![]() и положили
и положили ![]() . Коэффициенты уравнения зависят только от мембранного потенциала
. Коэффициенты уравнения зависят только от мембранного потенциала ![]() . Коэффициент
. Коэффициент ![]() называется максимальной калиевой проводимостью.
называется максимальной калиевой проводимостью.
Относительно проводимости ![]() для тока утечки А. Ходжкин и А. Хаксли предложили считать его постоянным:
для тока утечки А. Ходжкин и А. Хаксли предложили считать его постоянным: ![]() . В результате для описания процесса генерации потенциала действия была предложена система четырех уравнений, Первое из них -электрохимическое уравнение для баласа мембранных токов токов:
. В результате для описания процесса генерации потенциала действия была предложена система четырех уравнений, Первое из них -электрохимическое уравнение для баласа мембранных токов токов: ![]() . Напомним, что
. Напомним, что
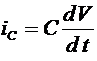 ,
, ![]() , (4)
, (4)
![]() ,
, ![]() . (5)
. (5)
Второе и третье уравнения описывают соответственно процессы открытия ![]() и закрытия
и закрытия ![]() -ворот для натриевых каналов. Наконец, четвертое уравнение отражает процесс открытия
-ворот для натриевых каналов. Наконец, четвертое уравнение отражает процесс открытия ![]() -ворот для калиевых каналов. Система уравнений Ходжкина -Хаксли имеет следующий вид:
-ворот для калиевых каналов. Система уравнений Ходжкина -Хаксли имеет следующий вид:
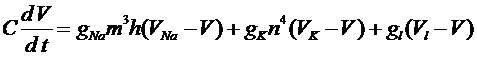 , (6)
, (6)
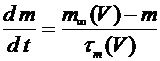 , (7)
, (7)
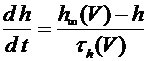 , (8)
, (8)
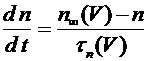 , (9)
, (9)
где, напомним, ![]() ,
, ![]() ,
, ![]() . В (7) -(9) функции
. В (7) -(9) функции ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() положительны и непрерывны. Функции
положительны и непрерывны. Функции ![]() ,
, ![]() ,
, ![]() относятся к классу так называемых сигмоидных функций, т.е. они монотонны обладают следующими свойствами:
относятся к классу так называемых сигмоидных функций, т.е. они монотонны обладают следующими свойствами: ![]() ,
, ![]() ,
, ![]() при
при ![]() и
и ![]() ,
, ![]() ,
, ![]() при
при ![]() . В работах А. Ходжкина и А. Хаксли указан их конкретный вид. Функции весьма сложно устроены. Реально система уравнений допускает только численное исследование. Ограничимся рядом замечаний на качественном уровне, иллюстрирующих впечатления от численного исследования. Входящие в систему (6) -(9) функции подобраны так, что выполнен ряд свойств. Она имеет устойчивое состояние равновесия
. В работах А. Ходжкина и А. Хаксли указан их конкретный вид. Функции весьма сложно устроены. Реально система уравнений допускает только численное исследование. Ограничимся рядом замечаний на качественном уровне, иллюстрирующих впечатления от численного исследования. Входящие в систему (6) -(9) функции подобраны так, что выполнен ряд свойств. Она имеет устойчивое состояние равновесия ![]() , где
, где ![]() ,
, ![]() -положительны. Пусть
-положительны. Пусть ![]() ,
, ![]() ,
, ![]() ,
, ![]() . Если значение
. Если значение ![]() не слишком велико, то в силу устойчивости состояния равновесия с течением времени
не слишком велико, то в силу устойчивости состояния равновесия с течением времени ![]() . Существует такое пороговое значение
. Существует такое пороговое значение ![]() , что при
, что при ![]() решения системы ведут себя по другому. Первоначально
решения системы ведут себя по другому. Первоначально ![]() убывает. Однако начинает расти величина
убывает. Однако начинает расти величина ![]() . Слагаемое
. Слагаемое ![]() уравнении (6) положительно, т.к.
уравнении (6) положительно, т.к. ![]() , а
, а ![]() близко к нулю. В результате производная
близко к нулю. В результате производная ![]() становится положительной. Мембранный потенциал начинает расти (момент начала генерации спайка). Вместе с ним растет и величина
становится положительной. Мембранный потенциал начинает расти (момент начала генерации спайка). Вместе с ним растет и величина ![]() , что приводит к дальнейшему увеличению
, что приводит к дальнейшему увеличению ![]() и ускорению роста
и ускорению роста ![]() . Однако, чуть позже начинает уменьшаться величина
. Однако, чуть позже начинает уменьшаться величина ![]() . Это замедляет рост
. Это замедляет рост ![]() , тем более, что уменьшается разность
, тем более, что уменьшается разность ![]() . Одновременно растет величина
. Одновременно растет величина ![]() . Соответствующее слагаемое
. Соответствующее слагаемое ![]() в уравнении (6) отрицательно, т.к.
в уравнении (6) отрицательно, т.к. ![]() и
и ![]() . Существует момент времени
. Существует момент времени ![]() , где
, где ![]() . Это точка максимума потенциала действия. Значение мембранного потенциала
. Это точка максимума потенциала действия. Значение мембранного потенциала ![]() относительно близко к
относительно близко к ![]() . После прохождения точки максимума значение производной
. После прохождения точки максимума значение производной ![]() определяется значением слагаемого
определяется значением слагаемого ![]() и становится отрицательным. Скорость убывания мембранного потенциала на нисходящем участке спайка меньше по абсолютной величине, чем скорость роста на восходящем участке. Нисходящий участок продолжительнее восходящего. В дальнейшем значение
и становится отрицательным. Скорость убывания мембранного потенциала на нисходящем участке спайка меньше по абсолютной величине, чем скорость роста на восходящем участке. Нисходящий участок продолжительнее восходящего. В дальнейшем значение ![]() становится отрицательным, а после этого
становится отрицательным, а после этого ![]() . Величины
. Величины ![]() ,
, ![]() ,
, ![]() также стремятся к своим равновесным значениям. Процесс генерации спайка закончен.
также стремятся к своим равновесным значениям. Процесс генерации спайка закончен.
Особо подчеркнем значимость факта, что система (6) -(7) обладает пороговыми свойствами. Существует такое критическое значение ![]() мембранного потенциала, что при начальном отклонении мембранного потенциала
мембранного потенциала, что при начальном отклонении мембранного потенциала ![]() с течением времени значение
с течением времени значение ![]() . Если же
. Если же ![]() , то через некоторое время (его называют скрытым, или латентным периодом) генерируется спайк. Мадель адекватно отражает пороговые свойства реального аксона. Если воздействовать на мембрану кратковременным толчком деполяризующего тока (отклоняющего мембранный потенциал в положительном направлении), то после слабого воздействия мембранный потенциал возвращается в равновесное состояние. Вследствие достаточно сильного воздействия спустя латентный период генерируется спайк.
, то через некоторое время (его называют скрытым, или латентным периодом) генерируется спайк. Мадель адекватно отражает пороговые свойства реального аксона. Если воздействовать на мембрану кратковременным толчком деполяризующего тока (отклоняющего мембранный потенциал в положительном направлении), то после слабого воздействия мембранный потенциал возвращается в равновесное состояние. Вследствие достаточно сильного воздействия спустя латентный период генерируется спайк.
Обсудим еще одно свойство аксона. Гигантские аксоны достигают длины нескольких десятков сантиметров. Мебранный потенциал на разных участках аксона может различаться. Аксон представляет собой так называемую распределенную систему. Если подействовать на некоторый участок аксона допороговым деполяризующим толчком тока, то мембранный потенциал на этом и некотором примыкаюшем к нему участке отклонится от равновесного состояния в положительном направлении. Такое отклонение называется локальным ответом. Возмущение с течением времени затухает, одновременно, как капля, растекаясь по аксону. В результате локальные ответы, возникшие при стимулировании различных участков, могут накладываться друг на друга, т.е., как говорят, суммироваться по пространству. Если же на участок аксона повторно подается деполяризирующий ток, то новый локальный ответ накладывается на предыдущий. Говорят, что происходит временное суммирование. Локальные ответы градуальны. Их амплитуда зависит от силы стимула (точнее, силы -длительности). В случае, когда локальный ответ в результате суммации, или же после достаточно сильного одноразового локального воздействия достигает порогового уровня, запускаются механизмы генерации потенциала действия. Зародившись локально, потенциал действия распространяется по аксону.
Явление распространения объясняется следующим образом. Цитоплазма - электролит, а, следовательно, хороший проводник. Если между точками существуют ненулевые разности потенциалов, то вдоль мембраны возникают токи (вблизи поверхности). Они направлены от точек с более высоким потенциалом к точкам, где он меньше, и подчиняются закону Ома. Вследствие этого потенциал на участках, прилегающих, к области генерации спайка, растет и достигает порогового значения. В результате эти участки сами генерируют потенциалы действия. Процесс распространяется вдель аксона.
Проведение нервных импульсов обладает важной особенностью. При “столкновении” они аннигилируют, т.е. уничтожаются. С помощью локального воздействия возбудим аксон в двух точках. По нему будут распространяться четыре импульса, два из которых движутся во встречных направлениях. Данные импульсы встретятся и не пропустят друг друга. Это связано с тем, что мембрана обладает следующим свойством. Во время потенциала действия и некоторое время после его завершения она не реагирует на воздействие. В частности, после завершения спайка некоторое время не способна сгенерировать нового потенциала действия. Данное состояние называется абсолютной рефрактерностью. Отметим, что постепенно абсолютная рефрактерность сменяется относительной. Для того, чтобы инициировать потенциал действия в состоянии относительной рефрактерности, нужно применить более мощный стимул, нежели в состоянии покоя.
Для описания процесса распространения потенциала действия вдоль аксона А. Ходжкин и А. Хксли модернизировали уравнение (6). Пусть ![]() -координата точки вдоль аксона, а
-координата точки вдоль аксона, а ![]() -значение мембранного потенциала в этой точке. Используется закон Ома, согласно которому ток через сечение пропорционален градиенту напряжения (в одномерном случае -производной). Ток через сечение
-значение мембранного потенциала в этой точке. Используется закон Ома, согласно которому ток через сечение пропорционален градиенту напряжения (в одномерном случае -производной). Ток через сечение ![]() равен
равен 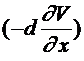 , где направление выбрано в сторону возрастания
, где направление выбрано в сторону возрастания ![]() , коэффициент
, коэффициент ![]() -проводимость. Выделим участок аксона, координаты точек которого заключены в интервале
-проводимость. Выделим участок аксона, координаты точек которого заключены в интервале ![]() . Суммарный ток через сечения
. Суммарный ток через сечения ![]() и
и ![]() равен
равен 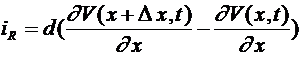 . Используя формулу Лагранжа получим:
. Используя формулу Лагранжа получим: 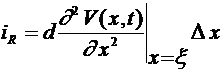 , где
, где ![]() . Через поверхность мембраны выбранного участка аксона проходят емкостной
. Через поверхность мембраны выбранного участка аксона проходят емкостной ![]() , натриевый
, натриевый ![]() , калиевый
, калиевый ![]() токи, а также ток утечки
токи, а также ток утечки ![]() . Пусть
. Пусть ![]() . Суммарный мембранный ток приближенно равен
. Суммарный мембранный ток приближенно равен ![]() . Привлекая закон Кирхгофа, получим:
. Привлекая закон Кирхгофа, получим: ![]() . Используя приведенные ранее формулы (4), (5) для мембранных токов и устремляя
. Используя приведенные ранее формулы (4), (5) для мембранных токов и устремляя ![]() к нулю, приходим к уравнению:
к нулю, приходим к уравнению:
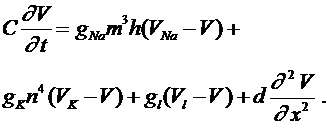 (10)
(10)
Его часто называют уравнением аксона Ходжкина -Хаксли. В (10) функции ![]() ,
, ![]() ,
, ![]() как и выше подчиняются уравнениям (7) -(9).
как и выше подчиняются уравнениям (7) -(9).
А. Ходжкин и А. Хаксли исследовали так называемые автоволновые решения системы уравнений, т. е. решения типа бегущих волн: ![]() (аналогично для
(аналогично для ![]() ,
, ![]() ,
, ![]() ). Здесь параметр
). Здесь параметр ![]() -скорость распространения волны. После подстановки в (11) для
-скорость распространения волны. После подстановки в (11) для ![]() получается обыкновенное дифференциальное уравнение:
получается обыкновенное дифференциальное уравнение:
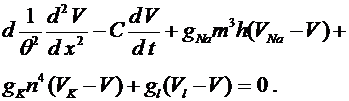 (11)
(11)
Система (7) -(9), (11) исследовалась численно. Методом подбора было найдено значение скорости ![]() , при которой система уравнений имела приемлемое по мнению авторов решение. Параметры полученного при исследовании системы уравнений распространяющегося потенциала действия оказались близкими к реально надлюдаемым в эксперименте.
, при которой система уравнений имела приемлемое по мнению авторов решение. Параметры полученного при исследовании системы уравнений распространяющегося потенциала действия оказались близкими к реально надлюдаемым в эксперименте.
Скорость распространения импульса по гигантскому аксону относительно не велика (по сравнению с аналогичными по назначению проводниками нервных импульсов у человека). Она растет с увеличением диаметра аксона. Увеличение диаметров аксонов избрала природа для высокоскоростного проведения импульсов у беспозвоночных. Однако, когда организму нужна не только быстрота реакции, но и огромное количество связей, гигантские аксона становятся не приемлимыми. Например, в зрительном нерве, где больше миллиона связей, для больших аксонов не хватило бы места. Длинные аксоны у человека и позвоночных животных покрыты достаточно толстым липидным слоем -миелиновой оболочкой (точнее, они обвиты клетками содержащими и вырабатывающими миелин). В миелиновой оболочке присутствуют регулярно расположенные разрывы, где мембрана аскона оголена. Разрывы называются перехватами Ранвье. Миелинизированные участки иеют длину порядка 1 -2 мм. Протяженность перехватов Ранвье около 1 мкм. Перехваты Равье способны генерировать спайки. Миелинизиро-ванные участки обладают емкостью и омическим сопротивлением, но они не генерируют спайков. Импульсы по миелинизированным участкам рапространяются пассивно, т.е. с затратами энергии на омическое сопротивление и на перезаряд участка, предствляющего собой конденсатор (обратимые потери). Суммарно затраты энергии не велики.
Рассмотрим миелинизированный участок, ограниченный двумя перехватами. Пусть левый перехват генерирует спайк. Возмущение (положительное) по миелинизированному участку передается правому перехвату. Когда отклонение мембранного потенциала правого участка достигает порогового значения, он генерирует спайк. Этот импульс не может вызвать генерацию нового спайка левым перехватом Ранвье, поскольку тот находится в рефрактерном состоянии (в состояниии невосприимчивости). Однако, данный перехват по той же схеме возбуждает перехват, следуюший за ним справа. По миелинизированному аксону, перескакивая от перехвата к перехвату, будет распространяться импульс. Процесс называется сальтаторным проведением возбуждения (saltare - прыгать). Оно обеспечивает скорость распространения импульсов в 20- 25 раз более высокую, чем в гигантских аксонах того же диаметра.
Описанный процесс сальтаторного проведения импульсов легко формализуется. В простейшем случае это выглядит следующим образом. Пусть ![]() и
и ![]() - мембранные потенцилы левого и правого перехвотов Ранвье, а
- мембранные потенцилы левого и правого перехвотов Ранвье, а ![]() -потенциал миелинизированного участка (считаем его во всех точках участка одинаковым). Тогда между участком и соответствующими перехватами проходят токи:
-потенциал миелинизированного участка (считаем его во всех точках участка одинаковым). Тогда между участком и соответствующими перехватами проходят токи: 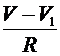 и
и 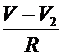 , где
, где ![]() -сопротивление перехода от перехвата к миелинизированному участку. Через миелиновую оболочку проходят емостной ток
-сопротивление перехода от перехвата к миелинизированному участку. Через миелиновую оболочку проходят емостной ток ![]() и ток утечки
и ток утечки ![]() , где
, где ![]() - суммарная емкость, а
- суммарная емкость, а ![]() суммарное сопротивление миелиновой оболочки. В силу первого закона Кирхгофа получаем:
суммарное сопротивление миелиновой оболочки. В силу первого закона Кирхгофа получаем:
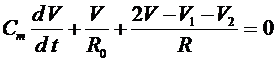 .
.
Для описания мембранных потенциалов ![]() и
и ![]() перехватов служит уравнение (6), в правую часть которого согласно закону Кирхгофа следует добавить для левого перехвата слагаемое
перехватов служит уравнение (6), в правую часть которого согласно закону Кирхгофа следует добавить для левого перехвата слагаемое 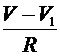 , а для правого -слагаемое
, а для правого -слагаемое 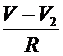 . Это токи, проходящие между соответствующими перехватами и миелинизированным участком. В результате получим уравнения:
. Это токи, проходящие между соответствующими перехватами и миелинизированным участком. В результате получим уравнения:
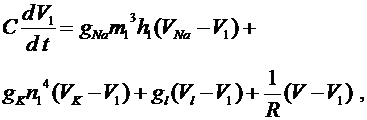
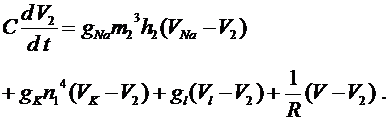
Величины ![]() ,
, ![]() и
и ![]()
![]() удовлетворяют уравнениям (7) -(9), в которых для левого и правого перехватов следует положить
удовлетворяют уравнениям (7) -(9), в которых для левого и правого перехватов следует положить ![]() и
и ![]() соответственно.
соответственно.
Сен 11 2018