
Статьи. Предметно
Первичная зрительная кора
Первичная зрительная кора
После того как в 1952 году была опубликована первая статья Куффлера о рецептивных полях ганглиозных клеток сетчатки с центром и периферией, открылись пути дальнейшей работы. Чтобы объяснить обнаруженные свойства этих клеток, нужны были дополнительные исследования на уровне сетчатки. Однако требовались также данные о следующих уровнях зрительной системы, чтобы понять, как мозг интерпретирует информацию, доставляемую глазами. Решение этих двух задач было сопряжено с огромными трудностями. При изучении центральных механизмов понадобилось несколько лет, чтобы разработать методику длительной (порядка нескольких часов) регистрации активности одиночной клетки. Еще труднее было подбирать зрительные стимулы, влияющие на эту активность.
Топографическое отображение
До того как стало возможным продолжение работ в этом направлении, уже имелись кое-какие сведения об участках мозга, связанных со зрительной функцией: достаточно хорошо были прослежены начальные этапы передачи зрительной информации (рис. 35). Мы знали, что волокна зрительного нерва образуют синапсы с клетками наружного коленчатого тела (НКТ) и что аксоны клеток НКТ оканчиваются в первичной зрительной коре. Было также ясно, что эти связи — от сетчатки к НКТ и от НКТ к коре — имеют топографическую организацию. Говоря о топографическом отображении, мы имеем в виду, что предшествующая структура проецируется на последующую упорядоченным образом: если идти вдоль какой-либо линии на сетчатке, то проекции последовательных точек этой линии в НКТ и в коре также образуют одну непрерывную линию. Таким образом, волокна зрительного нерва, выходящие из небольшого участка сетчатки, все будут направляться к какому-то небольшому участку НКТ, а все волокна от небольшой зоны НКТ придут в определенную зону зрительной коры. Такая организация связей не покажется удивительной, если мы вспомним упрощенную схему нервной системы на рис. 13: клетки группируются здесь в структуру, напоминающую стопку пластин, причем каждая клетка любой пластины получает входы от некоторой компактной группы клеток предыдущей пластины.
В сетчатке последовательные слои клеток расположены наподобие игральных карт в колоде, так что нервные волокна могут кратчайшим путем проходить с одного уровня на следующий. Клетки наружных коленчатых тел удалены на некоторое расстояние от клеток сетчатки, точно так же как кора удалена от НКТ и находится в другой части мозга. Тем не менее характер соединений между клетками остается тем же — проекции отдельных зон на более высокие уровни организованы так, как если бы эти зоны точно накладывались друг на друга.
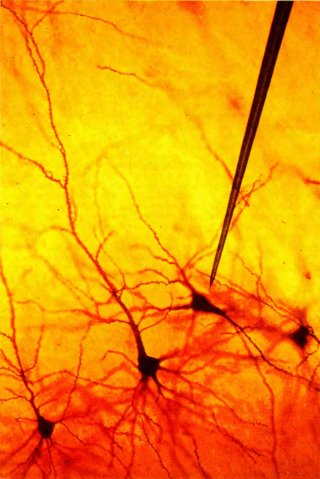
Рис. 34. В зрительной коре обезьяны окраска по Гольджи выявляет лишь несколько пирамидных клеток — ничтожную долю общего их числа в срезе. В действительности высота представленного здесь прямоугольного участка составляет около миллиметра. На микрофотографию наложено изображение (в том же масштабе) типичного вольфрамового электрода, применяемого при внеклеточной регистрации активности нейронов.
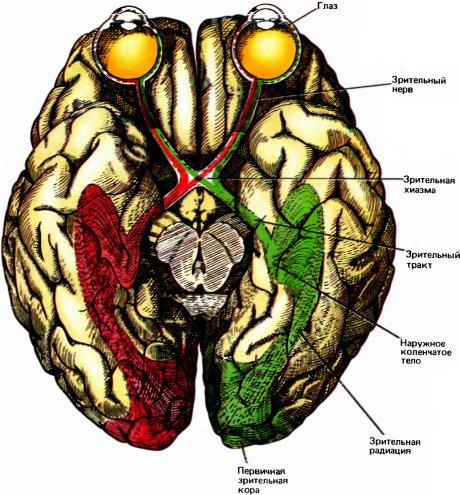
Рис. 35. Зрительные пути в мозгу человека, от глаз до первичной зрительной коры (вид снизу). Пусть входное изображение попадает на те половины обеих сетчаток, которые окрашены здесь в красный цвет (это их правые половины, поскольку мозг перевернут). На эти области проецируется противоположная половина пространства (левое зрительное поле); в конечном счете входная информация передается в правую половину мозга (пути ее передачи тоже окрашены в красный цвет). Это происходит потому, что примерно половина волокон, образующих зрительный нерв, переходит в хиазме на другую сторону, а другая половина остается на той же стороне. Таким образом, во-первых, каждое полушарие получает информацию от обоих глаз; во-вторых, каждое полушарие получает информацию о противоположной половине видимого мира.
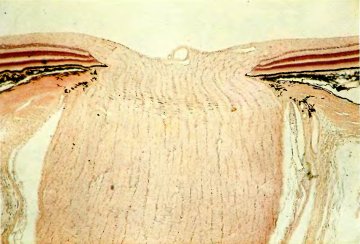
Рис. 36. Зрительный нерв в том месте, где он выходит из глаза, прерывая слои сетчатки, показанные слева и справа. Ширина области, представленной на микрофотографии, около 2 мм. Свободная зона вверху рисунка — это внутренняя среда глаза. На срезе видны слои сетчатки (сверху вниз): волокна зрительного нерва (светлые), три окрашенных слоя клеток и черный слой, содержащий пигмент меланин.
На выходе из глазного яблока волокна зрительного нерва просто собираются в пучок. Дойдя до НКТ, они расходятся и образуют своими окончаниями топографически упорядоченную проекцию. (Удивительно то, что в зрительном нерве на пути от сетчатки к НКТ эти волокна почти полностью перепутываются, но в НКТ снова «находят свои места».) Точно так же и волокна, выходящие из НКТ, расходятся в виде широкой полосы, которая идет через весь мозг к затылку и оканчивается в первичной зрительной коре, где проекция снова оказывается упорядоченной. После того как эти пути, пройдя через первичную зрительную кору и образовав синапсы в различных ее слоях, выходят из этой области и достигают других корковых зон, они опять образуют топографически упорядоченную проекцию. Поскольку на каждом уровне имеет место конвергенция связей, рецептивные поля в целом постепенно становятся все ближе, поэтому чем дальше от сетчатки, тем более размытым будет отображение зрительно воспринимаемой картины.
Еще одним важным и давним свидетельством в пользу топографической организации зрительных путей служат клинические наблюдения. Если поврежден определенный участок первичной зрительной коры, то развивается «локальная слепота», как если бы была разрушена соответствующая область сетчатки.
Итак, зрительный мир систематически отображен в структурах наружных коленчатых тел и коры. Однако в 50-х годах было неясно, что может означать такое отображение. В то время было еще не очевидно, что мозг обрабатывает получаемую информацию, преобразуя ее так, чтобы привести к более удобному для использования виду. Полагали, что зрительная сцена просто передается в мозг, а уж его задача — осмыслить ее (или эта задача, быть может, решается вообще не мозгом, а разумом). Из последующих глав мы узнаем, что такая нейронная структура, как первичная зрительная кора, производит глубокое преобразование поступающей информации. Нам почти ничего не известно о том, что происходит на дальнейших этапах ее переработки. Поэтому можно было бы сказать, что мы продвинулись не слишком далеко. Однако знание того, что определенная часть коры использует в своей работе вполне понятные принципы, дает основание для оптимизма — позволяет предполагать, что и остальные зоны коры работают так же. Возможно, настанет день, когда нам вообще не придется употреблять слово «разум».
Ответы клеток наружного коленчатого тела
Волокна, идущие в мозг от каждого глаза, проходят через зрительную хиазму (от названия греческой буквы «хи» — ?). В хиазме примерно половина волокон каждого зрительного нерва переходит на противоположную сторону мозга по отношению к данному глазу, а другая половина остается на той же стороне мозга. Пройдя хиазму, волокна направляются в несколько разных пунктов. Некоторые из них идут в нейронные структуры, имеющие отношение к таким специфическим реакциям, как движения глаз и зрачковый рефлекс. Однако большая часть волокон оканчивается в двух наружных коленчатых телах (НКТ). По сравнению с корой и множеством других отделов мозга эти тела устроены сравнительно просто — все или почти все из примерно полутора миллионов клеток в каждом НКТ имеют непосредственные входы от волокон зрительного нерва, и большинство клеток (но не все) посылают свои аксоны в кору мозга. Отсюда следует, что пути, проходящие через НКТ в кору, имеют только одно синаптическое переключение. Однако было бы ошибкой считать НКТ просто передаточной станцией. Сюда входят не только волокна из зрительного нерва, но и волокна, приходящие обратно из тех участков коры, на которые проецируются НКТ, а также из ретикулярной формации ствола мозга, имеющей отношение к процессам внимания и общей активации (arousal). Некоторые клетки НКТ имеют короткие аксоны (меньше миллиметра длиной), они не выходят за пределы НКТ, а образуют синаптические контакты с другими нейронами НКТ. Несмотря на эти усложнения, одиночные клетки НКТ отвечают на световые стимулы примерно так же, как и ганглиозные клетки сетчатки, и у них сходная структура рецептивных полей с on- и off-центрами и сходные ответы на цветовые стимулы. Таким образом, если говорить о переработке зрительной информации, НКТ, по-видимому, не производит никаких значительных преобразований сигналов. Что же касается незрительных входов НКТ и локальных синаптических связей, то пока мы просто ничего не знаем об их роли.
Представительство правой и левой сторон в зрительном пути
Волокна зрительного нерва распределяются между двумя НКТ не совсем обычным и на первый взгляд даже странным способом. Волокна от левой половины сетчатки левого глаза идут в НКТ той же стороны мозга, в то время как волокна от левой половины сетчатки правого глаза переходят в хиазме на другую сторону и, таким образом, попадают в то же левое НКТ (это показано на рис. 35). Аналогичным образом волокна от правых половин обеих сетчаток тоже оканчиваются в одном — правом — полушарии. Поскольку хрусталик создает на сетчатке перевернутое изображение, световые лучи, исходящие из правой половины зрительной сцены, проецируются на левые половины обеих сетчаток и информация передается в левое полушарие.
Термином зрительные поля обозначаются те участки окружающего мира или зрительной сцены, которые видны обоими глазами. Правое зрительное поле включает все точки, лежащие справа от вертикали, проходящей через ту точку, которую мы фиксируем взглядом (рис. 37). Важно отличать зрительные поля, т.е. то, что мы видим во внешнем мире, от рецептивного поля, которое соответствует тому участку внешнего мира, который «видит» одиночная клетка. Перефразируя то, что говорилось в начале этого раздела, можно сказать, что информация от правой половины поля зрения передается в левое полушарие.
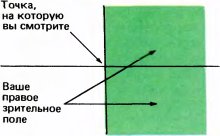
Рис. 37. Правое зрительное поле простирается вправо почти на 90°. В этом легко убедиться, если, быстро сгибая и разгибая палец, медленно смещать его по кругу вправо. Вверх поле зрения простирается на 60° или около того, вниз — примерно на 75°, а влево, по определению, доходит до вертикали, проходящей через точку, которую вы фиксируете взглядом.
Аналогичным образом устроены и многие другие отделы мозга. Например, осязательные и болевые сигналы от правой половины тела приходят в левое полушарие, а двигательное управление мышцами правой стороны тела осуществляется левым полушарием. Обширный инсульт левой половины мозга приводит к параличу и потере чувствительности правой половины лица, правой ноги и правой руки, а также к потере речи. Но, что менее известно, такой инсульт обычно сопровождается утратой зрения в правой половине зрительного поля, причем для обоих глаз. Чтобы установить наличие такой слепоты, невропатолог просит больного встать перед ним, закрыть один глаз и смотреть другим на кончик носа врача. Затем он начинает обследовать зрительные поля больного, перемещая в разные точки пространства свою руку или специальный зонд. В случае левостороннего инсульта можно показать, что больной ничего не видит справа от точки фиксации. Например, если врач поднимает в руке зонд и держит его между собой и больным немного выше своей головы, то при медленном движении руки справа налево (с точки зрения больного) больной ничего не видит до тех пор, пока белый рукав халата врача не пересечет среднюю линию и внезапно не появится в поле зрения больного. Если проверить другой глаз, то результат будет точно таким же. Полная правосторонняя гомонимная гемианопсия (так называют неврологи подобное выпадение половины поля зрения!) точно разделит и область центральной ямки (центр взора): если смотреть на слово was, фиксируя взглядом середину буквы а, то не будет видно буквы s, и даже от буквы а останется лишь ее левая половина — интересное, но весьма огорчительное переживание.
Такого рода тесты ясно показывают, что от каждого глаза сигналы передаются в оба полушария и что, наоборот, каждое полушарие мозга получает входные сигналы от обоих глаз. Это может показаться странным: после того что было сказано об осязательной и болевой чувствительности и о системе управления движениями, читатель может предположить, что левый глаз должен посылать сигналы в правое полушарие и наоборот. Однако в случае зрительной системы каждое полушарие мозга имеет дело не с противоположной стороной тела, а с противоположной половиной окружающего пространства. Но, впрочем, ситуация, когда сигналы от левого глаза передаются в основном в правое полушарие (и наоборот), имеет место у многих низших млекопитающих, таких как лошади и мыши, а у птиц и земноводных осуществляется даже полное перекрещивание зрительных нервов. У лошадей и мышей глаза расположены так, что они скорее направлены вбок, чем прямо вперед, в результате чего на большей части сетчатки правого глаза отображается правое зрительное поле, тогда как у приматов глаза направлены прямо вперед и на каждой сетчатке имеются отображения как правого, так и левого зрительного поля. Сделанное выше описание зрительных путей относится только к тем млекопитающим, у которых глаза смотрят более или менее прямо (как, например, у приматов) и поэтому видят почти один и тот же участок окружающего мира.
Сходным образом устроена и слуховая система. Понятно, что каждое ухо способно слышать звуки, исходящие как из левой, так и из правой половины окружающего пространства. Подобно глазам, каждое ухо передает информацию о звуке примерно в равной степени в обе половины мозга. При этом в слуховой системе, как и в зрительной, процесс передачи информации латерализован: звук, доходящий до каждого уха от некоторого источника с правой стороны, анализируется в стволе мозга путем сравнения амплитуд и моментов прихода сигналов к тому и другому уху, и в результате реакция на этот звук формируется в основном в высших отделах левой половины мозга.
Здесь мы говорим о начальных этапах переработки информации. Если справа от меня стоит человек, который словами или жестами побуждает меня сделать что-нибудь левой рукой, то сообщаемая им информация рано или поздно должна попасть в правое полушарие моего мозга. Однако первоначально сигналы должны прийти в слуховую или зрительную кору левого полушария, и только после этого они передаются в моторную кору правого полушария.
Между прочим, никто не знает, почему правая половина окружающего пространства обычно проецируется в левое полушарие головного мозга. Из этого правила есть одно важное исключение — полушария мозжечка (отдел мозга, который в основном управляет движениями) получают входные сигналы главным образом от той же самой, а не противоположной стороны окружающего пространства. Это усложняет работу мозга, поскольку все волокна, соединяющие одно полушарие мозжечка с моторной корой в другом полушарии большого мозга, должны переходить с одной стороны мозга на другую. По поводу такой организации можно пока сказать лишь то, что она кажется загадочной.
Слоистая структура наружного коленчатого тела (НКТ)
Каждое из НКТ содержит по шесть клеточных слоев. Отдельный слой имеет толщину в несколько клеток (от 4 до 10 и более). Весь этот шестислойный сэндвич согнут таким образом, что его поперечный срез имеет вид, показанный на рис. 38.
При переходе от сетчатки к НКТ простая схема, в которой каждый последующий слой клеток содержит проекцию предыдущего, становится более сложной. В НКТ проекции от сетчаток двух глаз объединяются, и два отдельных изображения, представленные на уровне ганглиозных клеток в сетчатках, проецируются на шесть слоев НКТ. Волокна от правого и левого глаза не конвергируют на одни и те же клетки НКТ — каждая из этих клеток получает сигналы только от какого-то одного глаза. Два множества клеток разнесены по отдельным слоям, так что в любом слое все клетки получают информацию только от одного глаза. Эти слои расположены таким образом, что проекции от правого и левого глаза чередуются. Так, в левом НКТ проекции располагаются в следующем порядке (от поверхности в глубину): левая, правая, левая, правая, правая, левая. Не совсем ясно, почему последовательность 5-го и 6-го слоев «перевернута» (иногда мне кажется, что это сделано для того, чтобы порядок проекций было труднее запомнить). У нас пока нет никакого вразумительного объяснения самого факта чередования проекций.
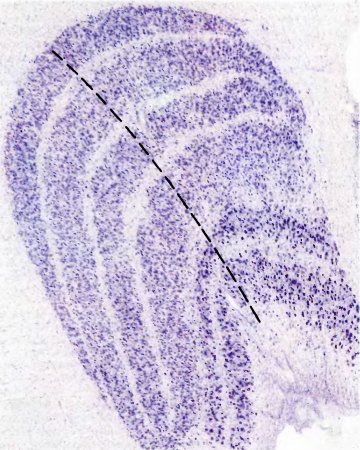
Рис. 38. Левое наружное коленчатое тело макака. Ясно видны шесть клеточных слоев. Срез сделан параллельно фронтальной плоскости; он специально окрашен для выявления тел нейронов (каждое из них выглядит как точка).
В целом шестислойная нейронная структура имеет одну общую топографию для всех слоев. Левые половины обеих сетчаток проецируются в слои левого НКТ (рис. 39), а правые половины — в слои правого НКТ. Любая точка в одном слое НКТ соответствует некоторой точке в поле зрения того или другого глаза. Если двигаться вдоль слоя НКТ, то соответственная точка в поле зрения будет перемещаться по некоторой траектории, определяемой характером отображения зрительного поля на НКТ. Если же двигаться перпендикулярно слоям НКТ (например, вдоль черной штриховой линии на рис. 38), как двигался бы микроэлектрод, проходя через разные слои, то соответствующие рецептивные поля клеток оставались бы в одном и том же участке поля зрения; при этом наблюдалось бы только чередование проекций от разных глаз, за исключением, конечно, того места, где идут подряд две проекции от одного глаза. Таким образом, каждая половина поля зрения шестикратно отображается на каждое из НКТ, трижды для каждого глаза, а проекции в слоях НКТ располагаются в точности друг под другом.
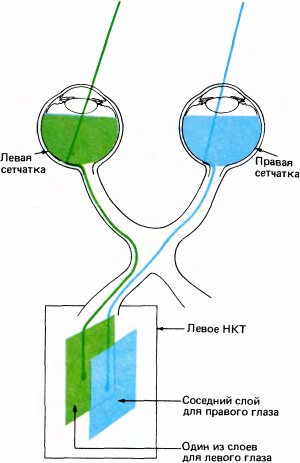
Рис. 39. При переходе от сетчатки к НКТ пространственная упорядоченность нейронов сохраняется, хотя на этом пути она временно исчезает, когда волокна собираются в пучок; в НКТ они снова «находят свои места».
Наружное коленчатое тело выглядит как состоящее из двух частей. Его подразделяют на вентральные, или нижние, слои и четыре дорсальных, или верхних, слоя (вентральный — расположенный ближе к брюшной стороне тела, дорсальный — к спинной стороне). Вентральная часть НКТ образует особую структуру, так как клетки в соответствующих слоях отличаются от клеток в остальных четырех слоях — они крупнее и по-иному отвечают на зрительные стимулы. В то же время четыре слоя дорсальной части НКТ как гистологически, так и по своим электрофизиологическим свойствам сходны друг с другом. Поскольку величина клеток в этих двух отделах различна, вентральные слои стали называть крупноклеточными, а дорсальные — мелкоклеточными.
Волокна, выходящие из шести слоев НКТ, объединяются в один широкий пучок, называемый зрительной радиацией, который идет вверх до первичной зрительной коры (см. рис. 35). Здесь эти волокна равномерно расходятся и перераспределяются так, что образуется целостная проекция с топографической организацией (это аналогично распределению волокон зрительного нерва при входе его в НКТ). И вот, наконец, мы подходим к коре.
Ответы клеток в коре
Главная тема этой главы — вопрос о том, как клетки в первичной зрительной коре отвечают на зрительные стимулы. Рецептивные поля нейронов НКТ имеют такую же организацию (разделение на центр и периферию), как и рецептивные поля ганглиозных клеток сетчатки, которые посылают свои аксоны к клеткам НКТ. Подобно ганглиозным клеткам сетчатки, нейроны НКТ различаются между собой главным образом свойствами рецептивного поля (on- или off-центр, местоположение в поле зрения) и особенностями ответов на цветовые стимулы. Возникает вопрос: а как обстоит дело с корковыми нейронами? Сходны ли они с клетками НКТ, посылающими в кору свои аксоны, или же у них появляются какие-то новые особенности? Ответ, как читатель уже должен догадаться, такой: корковые клетки действительно обладают новыми качествами, причем настолько необычными, что вплоть до 1958 года, когда их впервые стали изучать с помощью сложных световых стимулов, никто не мог даже приблизительно предсказать эти свойства.
Первичная зрительная кора (стриарная кора) представляет собой слой клеток толщиной 2 мм и площадью в несколько квадратных дюймов.[1] Для того чтобы дать представление о размерах этой нейронной структуры, можно привести такие цифры: если НКТ содержит полтора миллиона клеток, то стриарная кора — около 200 миллионов клеток. Анатомическая структура стриарной коры удивительно сложна, однако нет необходимости знать ее детали, чтобы понять, каким образом преобразуется здесь поступающая зрительная информация. Более подробно строение этого отдела будет рассмотрено в следующей главе, где будет обсуждаться вопрос о его функциональной архитектуре.
Как я уже говорил, процесс переработки информации в коре состоит из нескольких этапов. На первом этапе большинство клеток дает такие же ответы, как клетки НКТ. Рецептивные поля этих клеток обладают круговой симметрией. Это означает, что линия или граница (перепад освещенности) вызывает один и тот же ответ вне зависимости от ее ориентации. Регистрировать электрическую активность корковых клеток этого уровня непросто, так как они очень малы и расположены близко друг к другу. Пока еще не ясно, отличаются ли вообще ответы этих корковых клеток от ответов клеток НКТ (точно так же как не ясно, отличаются ли реакции клеток НКТ от ответов ганглиозных клеток сетчатки). Сложность гистологического строения НКТ и коры позволяет думать, что между ними должны быть какие-то различия и что их можно будет выявить, если знать, в чем их следует искать; однако узнать это может оказаться трудным делом.
Положение еще больше усложняется, когда мы переходим к ответам клеток на следующем корковом уровне. Вероятно, эти клетки должны получать входные сигналы от нейронов предыдущего уровня, обладающих рецептивными полями с центром и периферией. Вначале нам было совсем не просто выяснить, на какие зрительные стимулы отвечают эти клетки второго коркового уровня. В то время (конец 50-х годов) лишь очень немногие исследователи пытались регистрировать ответы одиночных нейронов зрительной коры. Те, кто это делал, получали противоречивые результаты. Они нашли, что клетки в зрительной коре работают, видимо, почти так же, как и в сетчатке, — были найдены как on-, так и off-клетки. Кроме того, был обнаружен еще один класс клеток, которые, казалось, вообще не отвечали на световые стимулы. Простота выявленных физиологических свойств корковых клеток на фоне просто дьявольской сложности морфологии коры ставила исследователей в тупик.
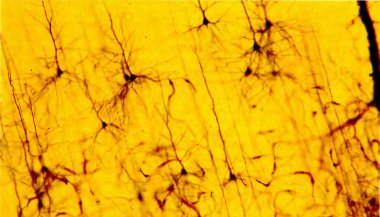
Рис. 40. Окрашенный по Гольджи срез первичной зрительной коры, на котором видно больше дюжины пирамидных клеток (но это лишь очень малая доля нейронов, содержащихся в срезе). Величина изображенного участка по вертикали — около 1 мм. (Темная полоса у правого края — кровеносный сосуд.)
Сегодня это очень просто объяснить. Во-первых, применялись неадекватные стимулы — для активации клеток коры по привычке просто освещали всю сетчатку рассеянным светом, хотя Куффлер еще за десять лет до того показал, что такая стимуляция даже для нейронов сетчатки далеко не оптимальна. Для большинства же корковых нейронов диффузный засвет сетчатки — не только не оптимальный, но вообще полностью неэффективный стимул. Если многие клетки наружных коленчатых тел еще реагируют, хотя и слабо, на диффузный засвет, то корковые клетки, даже относящиеся к первому корковому уровню и сходные с клетками НКТ, практически не отвечают на такую стимуляцию. Таким образом, первая приходящая в голову мысль, что для активации зрительных нейронов лучше всего стимулировать все рецепторы сетчатки, оказалась совершенно неверной. Во-вторых (и это еще парадоксальнее), оказалось, что те корковые клетки, которые давали on- или off-ответы, на самом деле были не клетками, а просто аксонами клеток НКТ. Настоящие корковые нейроны в этом случае вообще не отвечали на стимуляцию! Они считали ниже своего достоинства обращать внимание на такой примитивный стимул, как диффузный свет.
Именно так обстояло дело в 1958 году, когда Торстену Визелу и мне удалось осуществить одно из первых успешных отведений электрической активности коры у кошки. В этих опытах положение кончика микроэлектрода в корковой ткани было настолько стабильным, что можно было прослушивать активность одной и той же клетки в течение примерно девяти часов. Чтобы вызывать разряды клетки, мы использовали все возможные и невозможные средства — разве что сами не вставали на голову. (Как большинство корковых клеток, время от времени найденная клетка давала спонтанный разряд, но мы пытались убедить друг друга, что разряд был вызван именно нашими стимулами, и в результате потратили впустую много времени.) После нескольких часов безуспешных попыток у нас возникло смутное впечатление, что подача света на одну определенную область сетчатки вызывала некоторую реакцию, и мы сосредоточили свои усилия именно на этой области. В качестве стимулов мы в основном использовали круглые белые и черные пятнышки. Чтобы создать черное пятно, мы обычно брали предметное стекло размером 2,5?5 см, на которое был наклеен непрозрачный кружок; стекло вставлялось в специальное проекционное устройство, которое Сэмюэл Талбот сконструировал для подачи изображений на сетчатку. Для предъявления белых пятен мы брали медную пластинку такого же размера с небольшим отверстием (в те времена затраты на проведение исследований были гораздо меньшими). По прошествии примерно пяти часов напряженной работы нам неожиданно показалось, что, когда время от времени клетка случайно давала реакцию, эта реакция скорее была связана не с черным пятнышком, нанесенным на стекло, а с самим этим стеклом. В конце концов нам удалось установить, что реакцию клетки вызывала слабая, но четкая тень от края стеклянной пластинки, когда ее задвигали в окошко проекционного устройства. Вскоре мы убедились, что этот край работал как стимул только тогда, когда тень от него пересекала определенный участок сетчатки, причем эта тень должна была иметь вполне определенную ориентацию. Самым удивительным было резкое различие результатов в двух случаях — когда ориентация стимула была оптимальной (тогда разряд напоминал пулеметную очередь) и когда мы изменяли ориентацию стимула или производили тотальную засветку глаза яркой вспышкой (в этом случае никакой реакции не было).
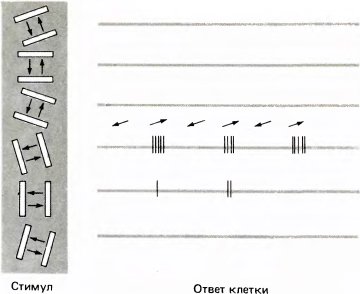
Рис. 41. Ответы одной из первых клеток, чувствительных к ориентации, которые Торстен Визел и я обнаружили в стриарной коре кошки в 1958 году. Данная клетка отвечала исключительно на стимул в виде движущейся щели с ориентацией, соответствующей положению часовой стрелки в 11 часов; при этом она реагировала на движение стимула вправо вверх и не реагировала на движение влево вниз.
Этот феномен был обнаружен самым первым и направил нас по ложному пути. Дело в том, что, как нарочно, обнаруженный тип клеток был как раз тот, который мы впоследствии назвали классом сложных клеток, а они относятся к уровню на две ступени выше, чем первый корковый уровень клеток с рецептивными полями, разделенными на центр и периферию. Хотя сложные клетки — наиболее распространенный тип клеток в стриарной коре, в их свойствах трудно разобраться, не познакомившись сначала с клетками предыдущего, промежуточного уровня.
Действительно, в отличие от клеток первого уровня, имеющих рецептивные поля с центром и периферией, у обезьяны клетки более высоких уровней проявляют совершенно иные реакции. Обычно точечные световые стимулы вызывают лишь слабую реакцию этих клеток или вообще неэффективны. Для того чтобы вызвать ответ клетки, сначала нужно найти надлежащий участок зрительного поля для предъявления стимула, т.е. найти соответствующий участок экрана, перед которым находится животное. Иными словами, сначала мы должны выявить рецептивное поле данной клетки. После этого выясняется, что наиболее эффективным стимулом для данной клетки служит линия, движущаяся в рецептивном поле в направлении, перпендикулярном ориентации линии. Такой линией могла быть узкая полоса света на темном фоне (щель), темная полоса на светлом фоне или же прямолинейная граница между темной и светлой областями. Некоторые клетки отдавали предпочтение, часто очень сильно выраженное, какому-то одному из этих трех стимулов, другие отвечали с примерно одинаковой интенсивностью на стимулы всех трех типов. Решающее значение имела ориентация линии — чаще всего клетка лучше всего реагировала на определенную оптимальную ориентацию, причем интенсивность ответа (число импульсов, возникавших при пересечении стимулом рецептивного поля) заметно снижалась при отклонении ориентации в любую сторону от оптимальной на 10–20 градусов; при еще большем отклонении реакция круто снижалась до нулевого уровня (см. рис. 41). Указанная цифра 10–20 градусов может показаться очень приблизительной. Вспомните, однако, что разница в положении часовой стрелки, показывающей «час» и «два», еще больше — 30 градусов. Когда ориентация стимула отличается от оптимальной на 90 градусов, типичная клетка, избирательно чувствительная к ориентации, перестает отвечать вообще.
В отличие от клеток на более низких уровнях зрительной системы нейроны, избирательно чувствительные к ориентации стимула, гораздо лучше отвечают на движущиеся, чем на неподвижные линии. Именно поэтому (см. рис. 41) при стимуляции таких нейронов мы использовали линии, движущиеся через рецептивное поле. Если использовать в качестве стимула неподвижную мелькающую (периодически вспыхивающую) линию, то зачастую клетка дает слабый ответ, и в этом случае предпочтительной оказывается такая же ориентация, как и при движущейся линии.
Многие клетки (вероятно, треть всей популяции) дают еще один характерный вид ответа на движущийся стимул. Вместо того чтобы давать один и тот же импульсный разряд независимо от направления движения, такие клетки отвечают более энергично при одном определенном направлении. Бывает даже так, что движение в одну сторону вызывает сильно выраженный ответ, а при движении в противоположную сторону нет вообще никакой реакции (это показано на рис. 41).
За один эксперимент можно оценить реакции 200–300 клеток, если после полного исследования одной клетки просто продвигать микроэлектрод дальше, до следующей клетки. Недостаток такой методики в том, что в одном прогоне можно исследовать только клетки, лежащие в коре на одной прямой линии: как только вы ввели тончайший микроэлектрод в корковую ткань, вы уже не можете сдвинуть его в поперечном направлении, не повредив при этом сам электрод или еще более нежную нервную ткань. С помощью такой методики регистрации самое большее, что мы можем сделать — это исследовать при одной проходке микроэлектрода примерно 50 клеток на миллиметр. Когда мы исследуем ориентационную избирательность нескольких сотен или тысяч клеток, оказывается, что все ориентации стимула встречаются примерно одинаково часто — вертикальная, горизонтальная и все промежуточные, наклонные ориентации. Если учесть характерные особенности окружающего нас мира, в который входят как деревья, так и линия горизонта, то возникнет вопрос: нет ли каких-нибудь выделенных ориентаций, таких как вертикаль и горизонталь, которые встречались бы чаще, чем другие? При попытках ответить на этот вопрос в разных лабораториях получали несколько разные результаты. Однако все исследователи соглашаются, что, если такие предпочтения действительно имеют место, они должны быть очень малы — настолько малы, что для выявления их требуется статистическая обработка данных. А в этом случае они вряд ли имеют какое-либо значение!
В стриарной коре обезьян примерно 70–80% клеток обладают свойствами ориентационной избирательности. Что касается кошек, то у них, по-видимому, все корковые клетки чувствительны к ориентации стимула, даже те, которые имеют прямые входы от наружных коленчатых тел.
Мы обнаружили заметные различия среди ориентационно-специфичных клеток, причем не столько в оптимальной ориентации стимула или в положении рецептивного поля на сетчатке, сколько в характере поведения клеток. Наиболее существенно различие между двумя классами нейронов — простыми и сложными клетками. Как можно догадаться по их названиям, клетки этих двух классов различаются по сложности своих ответных реакций. Поэтому мы сделали естественное предположение о том, что клетки с более простым поведением расположены в нейронной структуре коры ближе к ее входу.
Простые клетки
В большинстве случаев по ответам простых клеток на стимул в виде маленького светового пятна можно предсказать их реакцию на стимул сложной формы. Каждая из простых клеток, подобно ганглиозным клеткам сетчатки, клеткам НКТ и корковым клеткам с центрально-симметричными рецептивными полями, имеет небольшое четко очерченное рецептивное поле. Предъявление в пределах этого рецептивного поля стимула в виде светового пятнышка вызывает либо on-, либо off-реакцию в зависимости от того, в какой именно участок рецептивного поля подан стимул. Различие между простыми клетками и клетками предыдущих уровней заключается в конфигурации зон возбуждения и торможения. На предыдущих уровнях это центрально-симметричная конфигурация — имеется одна центральная on- или off-зона (возбуждающая или тормозная) и окружающая ее со всех сторон кольцевая зона с противоположными свойствами (тормозная или возбуждающая). Простые клетки коры более сложны. Зоны возбуждения и торможения в их рецептивных полях всегда разделены одной прямой линией или двумя параллельными линиями (рис. 42). Чаще всего встречается такая конфигурация, когда к длинной и узкой возбуждающей зоне с двух сторон примыкают более широкие тормозные зоны (рис. 42, А).
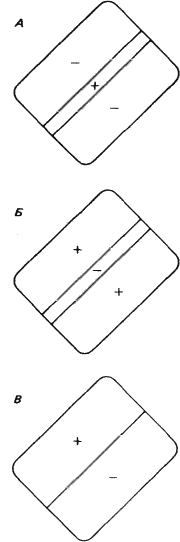
Рис. 42. Карты трех типичных рецептивных полей простых клеток. Оптимальными стимулами служили: для клетки А — светлая полоса против возбуждающей области (+); для клетки Б — темная линия, покрывающая тормозную зону (–); для клетки В — резкая граница «темное — светлое», совпадающая с границей между возбуждающей и тормозной зонами.
Для того чтобы проверить предполагаемую карту рецептивного поля, составленную путем тестирования маленьким световым пятном, мы попробовали использовать в качестве стимулов другие конфигурации. Вскоре мы выяснили, что чем большую долю той или иной зоны рецептивного поля покрывает данный стимул, тем сильнее выражено возбуждение клетки или ее торможение. Иными словами, имеет место пространственная суммация локальных воздействий. Мы обнаружили также явление антагонизма — взаимного погашения локальных воздействий при одновременной стимуляции возбуждающей и тормозной зоны. Таким образом, для клетки с рецептивным полем, изображенным на рис. 42, А, наиболее подходящим будет стимул в виде узкой полосы, расположенной в рецептивном поле так, чтобы она точно совпадала с возбуждающей зоной и не заходила в тормозную зону (см. рис. 43). Даже незначительное изменение ориентации этой полосы приведет к уменьшению эффективно действующей площади зоны возбуждения и затронет также тормозную зону; в результате частота разряда в ответе клетки уменьшится.
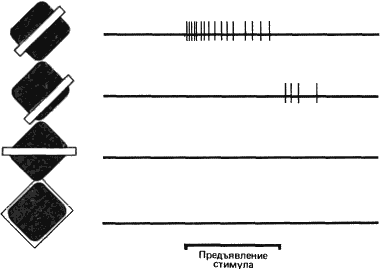
Рис. 43. Стимулы разных конфигураций вызывают различные реакции клетки с рецептивным полем такого типа, как А на рис. 42. Отрезком жирной линии внизу указан период (1 секунда), когда был включен стимул — светлая полоса. В первом случае (верхняя запись) показан ответ клетки на полосу оптимальных размеров, положения и ориентации. Во втором случае та же самая полоса покрывает только часть тормозной зоны (поскольку эта клетка не обладает спонтанной активностью, которая могла бы подавляться при торможении, здесь виден только разряд клетки при выключении стимула). В третьем случае полоса ориентирована так, что покрывает только малую часть возбуждающей зоны и соответственно малую часть тормозной зоны, и поэтому клетка не отвечает вообще. На нижней записи показан случай равномерного освещения всего рецептивного поля: ответа здесь тоже нет.
На рис. 42, Б и В показаны рецептивные поля простых клеток двух других типов, которые отвечают наилучшим образом на темные линии и на прямолинейные границы светлого и темного; при этом чувствительность клеток к ориентации стимула остается примерно такой же, как у клеток первого типа. Клетки всех трех типов совсем не реагируют на стимул в виде диффузного освещения. Такое взаимное погашение процессов возбуждения и торможения напоминает реакцию нейтрализации кислоты основанием, которую выполняют студенты в лабораторных работах по химии. Таким образом, уже на этом корковом уровне отмечается большое многообразие нейронов. Если взять класс простых клеток, то у них встречаются три или четыре разных типа рецептивных полей, причем имеются клетки, настроенные на любую из возможных ориентаций стимула, и клетки с рецептивным полем в любом участке поля зрения.
Величина рецептивных полей простых клеток зависит от их расстояния от центральной ямки. Однако в одной и той же зоне сетчатки тоже есть некоторые различия в размерах рецептивных полей. Самые маленькие рецептивные поля, расположенные в центральной ямке и около нее, имеют величину примерно 0,25?0,25°. Что касается клеток типа, показанного на рис. 42, А и Б, то ширина центральной зоны составляет у них не более нескольких угловых минут. Эта величина совпадает с минимальным диаметром рецептивных полей ганглиозных клеток сетчатки или клеток НКТ. Если же взять область далекой периферии, то размеры рецептивных полей простых клеток здесь могут достигать 1?1°.
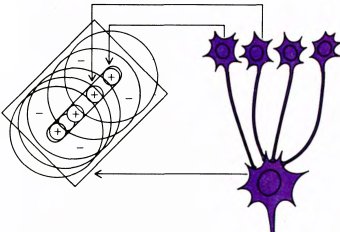
Рис. 44. Возможная схема связей, определяющих рецептивное поле простой клетки. Четыре клетки образуют возбуждающие синаптические связи с клеткой более высокого порядка. Каждая из клеток низшего порядка имеет рецептивное поле с радиальной симметрией, возбуждающим центром и тормозной периферией (это показано на схеме слева). Центры этих рецептивных полей лежат вдоль прямой линии. Если мы предположим, что с данной простой клеткой соединено много клеток, имеющих рецептивные поля с центром и периферией, причем центры этих рецептивных полей перекрываются и лежат на одной прямой, то рецептивное поле простой клетки будет состоять из длинной, узкой возбуждающей зоны и тормозных флангов. Избегая специальной терминологии, можно просто сказать, что небольшое светлое пятно в любом месте этого длинного и узкого прямоугольника будет приводить к сильному возбуждению одной или нескольких клеток с круглыми рецептивными полями и в результате — активировать, хотя и слабо, простую клетку. Если же стимулом будет длинная узкая полоса, способная активировать все клетки с круглыми рецептивными полями, то это приведет к сильной реакции простой клетки
Даже теперь, спустя двадцать лет, мы все еще не знаем, как устроены входные для корковых клеток цепи, от которых зависят специфические реакции этих клеток. Предложен ряд правдоподобных схем, и вполне может оказаться, что одна из этих схем или какая-то их комбинация окажется верной. Свойства простых клеток могут определяться нейронами предшествующего уровня с круглыми рецептивными полями; проще всего предположить, что простые клетки имеют прямые возбуждающие входы от многих клеток предыдущего уровня — таких, у которых центры рецептивных полей лежат в зрительном поле на одной прямой линии (рис. 44).
Несколько труднее предложить гипотетическую схему для клеток, избирательно реагирующих на границы темного и светлого (см. рис. 42, В). Возможен такой вариант: данная простая клетка имеет входы от двух наборов клеток предыдущего уровня, у которых центры рецептивных полей расположены с двух сторон от одной линии — по одну сторону клетки с on-центрами, а по другую с off-центрами, причем все эти входы возбуждающие. Во всех таких гипотетических схемах возбуждающий вход от клетки с on-центром по логике вещей эквивалентен тормозному входу от клетки с off-центром, при условии, что клетка с off-центром обладает спонтанной активностью.
Выяснение действительного механизма, определяющего реакции простых клеток, — задача непростая. Относительно данной простой клетки нужно будет узнать, от какого рода клеток она получает входные сигналы, например выяснить для каждой из предшествующих клеток структуру рецептивного поля, его местоположение и ориентацию (если она имеется), тип центра (on- или off-), а также характер посылаемых сигналов — возбуждающие они или тормозные. Поскольку методов получения таких сведений пока нет, мы вынуждены пользоваться косвенными подходами, а это увеличивает вероятность ошибки. Схема, представленная на рис. 44, кажется мне наиболее правдоподобной, так как она самая простая.
Сложные клетки
Сложные клетки соответствуют следующему уровню (или уровням) зрительного анализа. Они наиболее многочисленны в стриарной коре и составляют здесь, вероятно, около трех четвертей всей популяции нейронов. Первая из клеток, чувствительных к ориентации стимула, исследованная Визелом и мною, — та, которая отвечала на стимул в виде края стеклянного слайда, — почти наверное была сложной клеткой.
Общим свойством сложных и простых клеток является способность реагировать только на линии, ориентированные определенным образом. Сложные клетки, так же как и простые, отвечают на стимулы, предъявляемые в ограниченном участке поля зрения. От простых они отличаются тем, что реакции их нельзя объяснить формой и распределением возбуждающих и тормозных зон в рецептивном поле. Включение или выключение небольшого неподвижного пятна в пределах рецептивного поля редко вызывает ответ клетки. Даже на надлежащим образом ориентированную неподвижную полосу или границу клетка чаще всего не реагирует или дает лишь слабый, быстро затухающий ответ одного и того же типа как при включении, так и при выключении стимула. Однако, если должным образом ориентированная линия перемещается через рецептивное поле, возникает хорошо выраженный длительный разряд импульсов. Этот разряд начинается в момент, когда линия входит в рецептивное поле, и продолжается до тех пор, пока она не выйдет за его пределы (см. рис. 41, где показана схема ответа). Напротив, для того, чтобы вызвать длительный разряд простой клетки, нужно предъявить надлежащим образом ориентированную неподвижную линию в определенном участке рецептивного поля. Если же использовать движущуюся линию, то возникает только кратковременная реакция в тот момент, когда линия пересекает границу тормозной и возбуждающей зон, или же в то время, когда линия проходит через возбуждающую зону рецептивного поля. Те сложные клетки, которые способны реагировать на неподвижные световые «щели», полоски или границы, дают импульсный разряд независимо от того, в каком месте рецептивного поля расположен стимул, лишь бы ориентация его была подходящей. Однако те же стимулы совершенно неэффективны, если их ориентация далека от оптимальной (рис. 46).
Рис. 45. Активность этой клетки из слоя 5 стриарной коры кошки была записана в 1973 году с помощью внутриклеточного электрода Д. Эссеном и Дж. Келли из Гарвардской медицинской школы. Было проведено также картирование ее сложного рецептивного поля. Затем путем инъекции красителя проционового желтого выяснили, что это была пирамидная клетка.
Схемы на рис. 46 (для сложной клетки) и на рис. 43 (для простой клетки) демонстрируют существенное различие между двумя типами клеток: у простой клетки реакцию вызывает оптимально ориентированная линия лишь в очень узком диапазоне положений, а у сложной такая линия вызывает ответ, в каком бы участке рецептивного поля она ни предъявлялась. Это различие связано с существованием четко очерченных возбуждающих и тормозных зон в рецептивном поле простой клетки и с отсутствием их в рецептивном поле сложной клетки. Сложная клетка дает пример генерализации (нелокальности) ответа на линию в пределах более обширной области.
В целом у сложных клеток рецептивные поля несколько больше, чем у простых, но не намного. У макака в области центральной ямки чаще всего встречаются рецептивные поля сложных клеток величиной примерно 0,5?0,5°. В этой области сетчатки оптимальные размеры стимула как для простых, так и для сложных клеток составляют около двух угловых минут. Таким образом, «разрешающая способность» сложных клеток такая же, как у простых клеток.
Как и в отношении простых клеток, мы как следует не знаем, как организована система связей, передающая сигналы сложным клеткам. Однако и здесь нетрудно предложить несколько возможных схем их организации. Согласно простейшей из таких схем, сложная клетка получает входные сигналы от множества простых клеток, рецептивные поля которых имеют одну и ту же ориентацию, но размещены, частично перекрывая друг друга, по всему полю сложной клетки, как показано на рис. 47. Если связи от простых клеток к сложным возбуждающие, то всякий раз, когда в рецептивное поле сложной клетки попадает стимул в виде линии, возбуждаются некоторые простые клетки. В результате будет возбуждаться и сложная клетка.
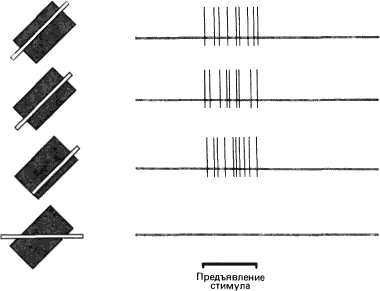
Рис. 46. Длинная и узкая полоса света вызывает реакцию сложной клетки независимо от того, в каком месте рецептивного поля она предъявлена, если только ее ориентация оптимальна (три верхние записи). Если ориентация полосы отличается от оптимальной, клетка реагирует слабее или не отвечает вовсе (нижняя запись).
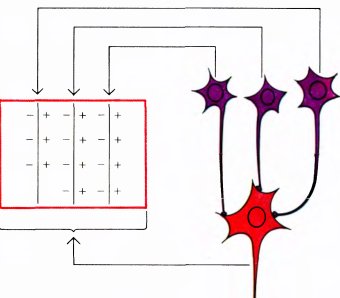
Рис. 47. Схема связей, которая позволила бы объяснить наблюдаемые свойства сложной клетки. Мы предполагаем, что (как и на рис. 44) к одной сложной клетке могут приходить возбуждающие сигналы от большого числа простых клеток (здесь показаны только три). Каждая простая клетка наилучшим образом отвечает на вертикальную границу между светлым (слева) и темным (справа) участками. Предполагается, что рецептивные поля простых клеток разбросаны в пределах прямоугольника и перекрываются. Если стимул в виде такой границы подается в любое место прямоугольника, то некоторое число простых клеток активируется и это в свою очередь вызывает ответ сложной клетки. Из-за эффекта адаптации синапсов только движущийся стимул будет вызывать непрерывное возбуждение сложной клетки.
Обычно в ответ на неподвижную линию сложная клетка дает короткий импульсный разряд (даже если стимул остается включенным). В этом случае мы говорим, что происходит адаптация ответа. Если же перемещать линию в рецептивном поле сложной клетки, наблюдается непрерывный разряд: адаптация преодолевается в результате последовательного срабатывания новых простых клеток.
Читатель, должно быть, заметил, что обе приведенные схемы связей — от клеток с круглыми рецептивными полями к простым клеткам (рис. 44) и от простых клеток к сложным (рис. 47) — предполагают использование возбуждающих связей. Однако в этих двух случаях процессы возбуждения должны быть совершенно различными. В первой из этих схем требуется суммирование одновременных сигналов от клеток с круглыми рецептивными полями, лежащих на одной линии. Во второй же схеме для активации сложной клетки движущимся стимулом необходимо последовательное возбуждение множества простых клеток. Было бы интересно выяснить, с какими морфологическими различиями (если они есть) связано это различие в механизме суммации.
Дирекциональная избирательность
Многие сложные клетки лучше реагируют на движение стимула в одном направлении, чем в противоположном. Различие в реакции часто бывает весьма резким — при одном направлении движения возникает энергичный ответ, а при обратном направлении клетка вообще не отвечает (рис. 48). Как выяснилось, примерно 10–20% клеток в верхних слоях стриарной коры проявляют заметную дирекциональную избирательность. Остальные клетки, по-видимому, не обладают такой избирательностью — мы тщательно исследовали ответы клеток с помощью ЭВМ, пытаясь обнаружить хотя бы небольшое различие в ответах на движение стимула в противоположных направлениях. Таким образом, существуют, видимо, два разных класса клеток — один из них четко проявляет дирекциональную избирательность, другому она не свойственна.
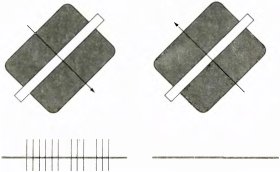
Рис. 48. Реакции данной сложной клетки на движение оптимально ориентированной полосы в противоположных направлениях различны. Длительность каждой записи около 2 секунд. (Для таких клеток не особенно важно, насколько быстро движется полоса; обычно клетка не отвечает только при очень быстром движении, когда полоса кажется размытой, или же при таком медленном, что его вообще трудно заметить.)
Если прослушивать импульсную реакцию клетки с сильно выраженной дирекциональной избирательностью, то создается впечатление, что при движении линии в одном направлении стимул как бы резко подталкивает клетку и заставляет ее разряжаться, а при движении в противоположном направлении происходит как бы сбой и стимул становится неэффективным (это напоминает ситуацию, когда вы, заводя часы, поворачиваете головку заводного механизма назад и раздается характерный треск).
Мы не знаем, как устроены входные сети таких клеток с дирекциональной избирательностью. Возможно, что на вход такой клетки подключены простые клетки, реакции которых на движение стимула в противоположных направлениях неравнозначны, асимметричны. Рецептивные поля таких простых клеток асимметричны, как, например, поле, представленное на рис. 42, В. Другую возможную схему предложили в 1965 году Г. Барлоу и У. Левик для объяснения дирекциональной избирательности некоторых клеток в сетчатке кролика — клеток, которых, по-видимому, нет у обезьяны. Если применить их схему к сложным клеткам коры, то нужно предположить наличие промежуточного слоя клеток, расположенных между простыми и сложными клетками (как на рис. 49). Представим себе, что клетка из промежуточного слоя имеет возбуждающий вход от одной простой клетки и тормозный вход — от другой клетки, рецептивное поле которой непосредственно примыкает к предыдущему, и притом всегда с одной стороны. Предположим далее, что тормозный путь включает некоторую задержку, возможно, за счет включения еще одной промежуточной клетки. В таком случае, если стимул движется в одном направлении, скажем справа налево (как на рис. 49, иллюстрирующем модель Барлоу и Левика), то промежуточная клетка возбуждается сигналом с одного из своих входов как раз в тот момент, когда действует торможение, приходящее от другой клетки, рецептивное поле которой только что пересек стимул. Возбуждающий и тормозный эффекты нейтрализуют друг друга, и в результате клетка не дает импульсного разряда. Если же стимул движется в обратном направлении, торможение возникает слишком поздно для того, чтобы предотвратить импульсный разряд. Если множество таких промежуточных клеток конвергирует на какую-то клетку, расположенную на следующем, третьем, уровне, то эта клетка будет обладать свойствами сложной клетки с дирекциональной избирательностью.
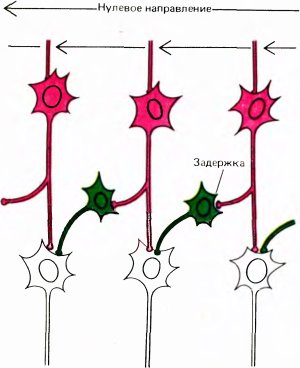
Рис. 49. Эту схему предложили X. Барлоу и У. Левик для объяснения свойства дирекциональной чувствительности. Синапсы, которые красные клетки образуют на зеленых клетках, — возбуждающие, а синапсы, образуемые на белых клетках, — тормозные. Мы предполагаем, что три белые клетки (внизу) конвергируют на одну «главную» клетку.
У нас нет прямых доказательств правильности тех или иных иерархических схем, объясняющих наблюдаемые реакции клеток, т.е. схем, в которых клетки на каждом следующем уровне организуются на основе элементов предыдущего уровня. Тем не менее у нас есть веские основания считать, что нервная система организована по иерархическому принципу. Одно из самых серьезных оснований для такого заключения связано с морфологией — например, у кошек простые клетки сосредоточены в четвертом слое стриарной коры, т.е. в том слое, который получает входы от клеток наружных коленчатых тел, тогда как сложные клетки располагаются в слоях коры выше и ниже — на одно или два синаптических переключения дальше от входа. Таким образом, хотя мы не можем дать точную схему связей на каждом уровне, у нас есть достаточные основания полагать, что такая схема существует.
Главная причина того, что в сложных клетках мы видим результат определенной организации клеток с круглыми рецептивными полями с центром и периферией, — это очевидная необходимость производить переработку информации логически в два этапа. Я хотел бы подчеркнуть слово логически, поскольку в целом нужное преобразование, по-видимому, могло бы быть физически осуществлено и в один этап — если суммировать на отдельных разветвлениях дендритов сложных клеток входные сигналы от клеток с круглыми рецептивными полями. В этом случае каждое разветвление выполняло бы функции отдельной простой клетки, посылая к телу клетки (и соответственно к аксону) сигналы электротоническим путем (с помощью пассивного электрического процесса) всякий раз, когда какая-то линия попадет в определенную зону соответствующего рецептивного поля. В этом случае такая клетка сама проявляла бы свойства сложной клетки. Однако само наличие простых клеток заставляет думать, что мы не должны строить таких сложных воображаемых конструкций.
Значение клеток, чувствительных к движению (включая некоторые комментарии к тому, как мы «видим»)
Почему столь многочисленны клетки, чувствительные к движению? Прежде всего приходит в голову, что такие клетки сигнализируют о том, имеется ли в поле зрения движущийся объект. Для животных, в том числе и для нас с вами, изменения внешнего мира гораздо важнее, чем подтверждения его неизменности, идет ли речь о выживании хищника или жертвы. Поэтому не удивительно, что большинство корковых клеток лучше реагирует на движущиеся объекты. Следуя этой логике, вы должны теперь усомниться — как же тогда мы проводим анализ неподвижных зрительных сцен, если в интересах чувствительности к движению так много клеток, настроенных на определенную ориентацию стимула, нечувствительно к неподвижным контурам. Чтобы получить ответ на этот вопрос, нужно сделать небольшое отступление и остановиться на некоторых важных, но противоречащих интуиции данных относительно того, как мы «видим».
Прежде всего естественно было бы ожидать, что при обследовании окружающего мира наши глаза будут плавно сканировать всю сцену непрерывными движениями. На самом же деле при фиксации какого-то объекта происходит вот что: сначала мы устанавливаем глаза так, чтобы изображение этого объекта попало в область центральной ямки обоих глаз, затем мы удерживаем глаза в таком положении в течение короткого времени, скажем полсекунды, потом глаза скачком перемещаются в новую позицию и фиксируют новую мишень, которая находится где-то в другом месте зрительного поля и привлекает к себе внимание тем, что несколько сдвигается относительно фона или имеет какую-то интересную форму. Во время такого скачка, или саккады, скорость движения глаз столь велика, что зрительная система не успевает отреагировать на перемещение изображения по сетчатке и мы его просто не замечаем. (Возможно, что в некотором смысле зрение отключается на период скачка с помощью какой-то сложной нейронной схемы, которая связывает глазодвигательные центры с главным зрительным путем.) Таким образом, процесс обследования зрительного поля при чтении или просто при рассматривании окружающего пространства состоит из серии быстрых скачков с одной точки на другую.
Специальная запись движений глаз позволяет наглядно продемонстрировать, насколько незаметны для нас эти скачки. Чтобы записывать движения глаз, берут крошечное зеркальце и прикрепляют его к контактной линзе сбоку — так, чтобы оно не мешало видеть. После этого на зеркальце направляют пучок света, который, отражаясь, создает на экране небольшое световое пятнышко. Есть еще один способ регистрации движений глаз, более современный; он разработан Д. Робинсоном в Институте Уилмера (университета Джонса Гопкинса). При этой методике по краю контактной линзы на глазу испытуемого монтируется крохотная катушка индуктивности, а самого испытуемого помещают между двумя взаимно перпендикулярными обручами величиной с велосипедное колесо, на которых расположены другие катушки индуктивности. Протекание тока в этих катушках ведет к появлению тока в катушке на контактной линзе. После калибровки такой системы можно регистрировать движения глаз с большой точностью. Для самого бедняги испытуемого ни тот, ни другой способ регистрации движений глаз нельзя назвать приятным!
В 1957 году советский психофизик А. Л. Ярбус записал движения глаз испытуемых при рассматривании различных изображений, в частности таких, как сцена в лесу и женское лицо (рис. 50). На этих записях периоды фиксации взора представлены точками, а точки соединены линиями, показывающими траекторию движения глаз во время скачков. Даже беглое ознакомление с этими удивительными записями дает много сведений о работе нашей зрительной системы — даже о том, какие предметы и детали окружающего мира нас больше интересуют.
Итак, первый факт, противоречащий нашей интуиции, состоит в том, что при зрительном осмотре глаза перескакивают с одной интересной точки на другую и что нельзя осматривать неподвижную сцену плавными движениями глаз. Задача глазодвигательной системы, видимо, заключается не в том, чтобы удерживать изображение на сетчатках неподвижным, а в том, чтобы предотвращать его плавное смещение. Если же движется вся зрительная сцена, как бывает, когда мы смотрим из окна поезда, то мы прослеживаем эту сцену, фиксируя взором какой-нибудь объект и поддерживая его фиксацию путем плавного движения глаз до тех пор, пока объект не выйдет из зоны видимости, после чего делаем скачок и фиксируем новый объект. Такая последовательность движений глаз — плавное прослеживающее движение, скажем вправо, а затем саккада влево — называется нистагмом. Вы сможете сами увидеть такого рода движения, когда окажетесь в поезде или в трамвае: понаблюдайте за движениями глаз ваших попутчиков, когда они будут рассматривать через окно окружающий пейзаж (будьте только осторожны, чтобы ваше внимание не было неверно истолковано!). Управление скачками глаз при рассматривании интересных элементов сцены с целью перевода их изображений в центральную ямку осуществляется из верхних бугорков четверохолмия. Это показал в 1978 году в серии впечатляющих работ П. Шиллер из Массачусетсского технологического института.

Рис. 50. Испытуемый рассматривает картину, и в это время у него все время регистрируется положение глаз и, следовательно, направление взгляда. Глаза совершают скачок и тотчас останавливаются (в этот момент в записи появляется небольшая точка), затем следует скачок к новому интересному месту. Создается впечатление, что глазу трудно перескочить в те места картины, где нет резких перепадов яркости.
Вторая группа фактов относительно того, как мы видим, еще больше противоречит нашему субъективному впечатлению. Когда мы рассматриваем неподвижную сцену и наш взгляд фиксирует какую-то точку, привлекающую внимание, то эта фиксация не бывает абсолютно неподвижной. Несмотря на все наши попытки жестко фиксировать точку, глаза не остаются в полном покое, а совершают непрерывные микродвижения, называемые микросаккадами. Они совершаются несколько раз в секунду и направлены более или менее случайно, достигая амплитуды 1–2 угловых минут. В 1952 году Л. Риггс и Ф. Рэтлифф из Университета Брауна и Р. Дитчберн и Б. Гинсборг из Редингского университета одновременно и независимо обнаружили, что если изображение на сетчатке искусственно стабилизировать (специальными методами), устранив его смещения относительно сетчатки, то зрительный образ спустя примерно секунду как бы «выцветает» и поле зрения становится совершенно пустым! (Простейший способ стабилизации состоит в том, что точечный источник света прикрепляют к контактной линзе; когда глаза движутся, источник света тоже движется и световое пятно быстро становится невидимым.) Если после стабилизации хотя бы чуть-чуть сместить изображение на сетчатке, световое пятно тотчас появляется снова. Очевидно, микросаккады необходимы для того, чтобы непрерывно видеть неподвижные объекты. Как будто бы Природа, создавая зрительную систему, особенно заботилась о восприятии движения и поэтому постаралась обеспечить нечувствительность клеток к неподвижным объектам, однако потом ей пришлось изобрести микросаккады, для того чтобы сделать и неподвижные объекты видимыми.
Можно предположить, что в этом процессе участвуют сложные клетки коры, особо чувствительные к движению стимула, но, вероятно, не участвуют клетки с дирекциональной избирательностью, так как микросаккады явно случайно распределяются по направлениям. С другой стороны, механизм дирекциональной избирательности должен, по-видимому, быть полезным для выявления движений объектов относительно неподвижного фона, сигнализируя о наличии движения и о его направлении. Для того чтобы следить за движущимся объектом на неподвижном фоне, нужно фиксировать объект и перемещать взор вместе с ним. В этом случае изображение всего остального будет передвигаться по сетчатке (такая ситуация в других случаях встречается редко). Перемещение всех деталей неподвижного фона по сетчатке должно приводить к бурной активности клеток коры.
Концы линий как зрительные стимулы
В стриарной коре встречается еще один вид клеток. Обычно для простых и сложных клеток характерна пространственная суммация — чем длиннее стимульная линия, тем лучше ответ. Однако реакция усиливается лишь до тех пор, пока длина линии не достигнет размеров рецептивного поля: дальнейшее удлинение линии не приводит к более энергичному ответу. В отличие от этого у клеток, реагирующих на концы линий (end stopped cells), удлинение линии до известного предела продолжает улучшать ответ, а если линия выходит за этот предел (в одном или в обоих направлениях), то ответ ослабевает (рис. 51, Б). Некоторые клетки, которые мы называем «реагирующими исключительно на конец линии» (completely end stopped cells), вообще не отвечают на предъявление стимула в виде длинной линии. Ту зону, с которой можно вызвать ответ клетки, мы называем зоной активации (или возбуждающей зоной), а зоны, расположенные с одного или с двух концов, — зонами торможения (или тормозными зонами). Таким образом, все рецептивное поле подобной клетки состоит из возбуждающей зоны и тормозной зоны (или зон) по краям. Стимул оптимальной ориентации, активирующий клетку с возбуждающей зоны, вызывает максимальное торможение за пределами этой зоны (с одной или с двух сторон). Это можно показать путем повторной стимуляции возбуждающей зоны линией оптимальной длины с оптимальной ориентацией при одновременном тестировании внешней зоны линиями разной ориентации (как показано на рис. 52).
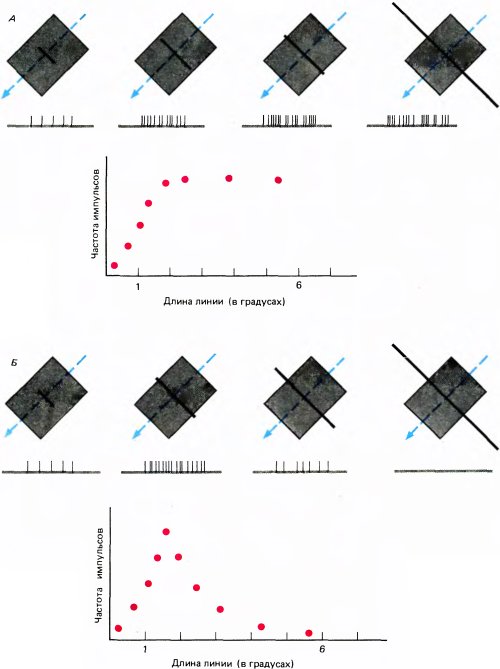
Рис. 51. А. Ответ обычной сложной клетки на светлую линию различной длины. Длительность каждой записи — 2 секунды. Как показывает график зависимости ответа от длины линии, реакция данной клетки усиливается, пока линия не достигает длины около 2°, после чего ответ остается неизменным. Б. Для этой клетки, реагирующей на концы линий, ответ возрастает, пока длина линии не достигнет 2°, а затем снижается, так что линия длиной 6° не вызывает никакого ответа.

Рис. 52. Для данной клетки, реагирующей на концы линий, предъявление в средней возбуждающей зоне одной полосы с оптимальной ориентацией приводит к возникновению энергичного ответа. Вовлечение в стимуляцию одной из тормозных зон почти полностью подавляет реакцию клетки; однако, если в этой тормозной зоне стимул имеет иную ориентацию, первоначальный ответ клетки не изменяется. Таким образом, и для возбуждающей зоны, и для тормозных зон оптимальная ориентация одинакова.
Вначале мы думали, что такие клетки относятся к следующему уровню иерархии корковых нейронов, на уровень выше сложных клеток. Согласно простейшей схеме возможной организации таких клеток, они могли бы иметь один или несколько возбуждающих входов от обычных сложных клеток с рецептивными полями, находящимися в возбуждающей зоне, и тормозные входы от сложных клеток с такой же ориентацией рецептивного поля, расположенных вне зоны возбуждения (эту схему поясняет рис. 53). Согласно другой возможной схеме (рис. 54), клетка имеет возбуждающий вход от клеток с небольшим рецептивным полем (а) и тормозный вход от клеток с большим рецептивным полем (б). Предполагается, что клетки, подающие тормозные сигналы, максимально чувствительны к длинным линиям, но слабо возбуждаются короткими линиями. Эта вторая схема (аналогичная модели клеток, имеющих рецептивные поля с центром и периферией, см. стр. 60) — одна из немногих схем, получивших частичное подтверждение. Чарлз Гилберт из Рокфеллеровского университета в Нью-Йорке установил, что сложные клетки в слое 6 стриарной коры обезьяны имеют как раз те свойства, которые нужны для торможения, предполагаемого в данной схеме. Он также показал, что если инактивировать эти клетки путем локальных инъекций, то расположенные в верхних слоях коры клетки, реагирующие на концы линий, теряют способность отвечать торможением на конец линии.
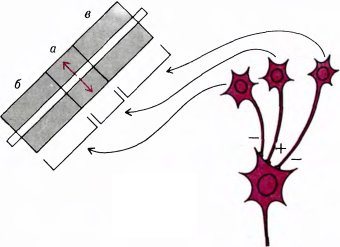
Рис. 53. Одна из схем, позволяющих объяснить поведение сложной клетки, реагирующей на концы линий. На такую клетку конвергируют выходы трех обычных сложных клеток; при этом у одной из этих клеток рецептивное поле совпадает по положению с возбуждающей зоной клетки, реагирующей на конец стимула (а), а у двух других клеток рецептивные поля лежат по обе стороны от возбуждающей зоны (б и в). Первая клетка образует на выходе возбуждающий синапс (+), а две другие — тормозные синапсы (–).
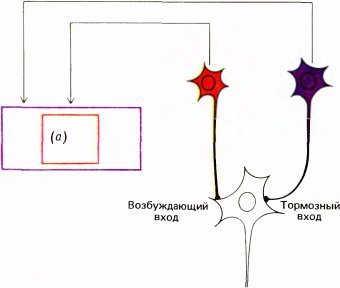
Рис. 54. Альтернативная схема, в которой торможение осуществляет одна клетка с рецептивным полем, соответствующим всей зоне а+б+в на рис. 53. Для того чтобы эта схема работала, мы должны предположить, что тормозная клетка слабо реагирует на короткую линию при стимуляции зоны а, но интенсивно отвечает на длинную линию.
Уже после того как были предложены описанные выше модели, Дж. Генри (Канберра, Австралия) обнаружил простые клетки, реагирующие на конец линии, схема рецептивных полей которых представлена на рис. 55. Схема организации подобных клеток аналогична предложенной выше первой схеме, за исключением того, что входы здесь не от сложных клеток, а от простых. Таким образом, ответы сложных клеток, реагирующих на концы линий, могли бы определяться сочетанием возбуждающего входа от одного набора сложных клеток и тормозного входа от другого набора (как в схемах, представленных на рис. 53 и 54) или же конвергенцией входов от нескольких простых клеток, реагирующих на концы линий.
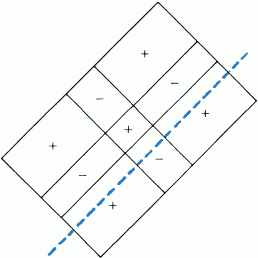
Рис. 55. Предполагается, что простая клетка, реагирующая на концы линий, может быть организована за счет конвергенции входных связей от трех обычных простых клеток. (Одна из них, имеющая on-центр, могла бы оказывать возбуждающее действие; две другие могли бы иметь on-центры и возбуждающие выходы или же off-центры и тормозящие выходы.) Вместо этого входные сигналы могли бы поступать непосредственно от клеток с круглыми полями (с центром и периферией) при использовании более сложного варианта схемы, показанной на рис. 44.
Оптимальным стимулом для клетки, настроенной на конец линии, служит отрезок линии определенной длины. Для такой клетки, которая реагирует на границу, причем с одной стороны отвечает только на ее конец, идеальным стимулом будет угол; для клетки, реагирующей на оба конца светлых или темных линий, наилучшим стимулом окажется короткая белая или черная линия, а также такая кривая линия, которая попадает в возбуждающую зону и не попадает из-за своей кривизны в тормозную зону (так будет, если ориентация ее концов отличается не менее чем на 20–30° от ориентации ее средней части; подобный случай показан на рис. 56). Таким образом, клетки, реагирующие на концы, можно рассматривать как клетки, чувствительные к углам, к кривизне или к резким изломам линий.
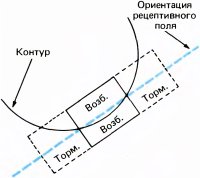
Рис. 56. Для клетки, реагирующей на концы линий (вроде представленной на рис. 52), эффективным стимулом может служить искривленная граница.
Нейрофизиология одиночных клеток и зрительное восприятие
Сам по себе факт, что какая-то клетка мозга реагирует на зрительные стимулы, еще не означает, что она непосредственно участвует в восприятии. Например, многие нейронные структуры в стволе мозга, относящиеся главным образом к зрительной системе, предназначены только для вспомогательных функций — для управления движениями глаз или сужением зрачка, для фокусировки изображения с помощью хрусталика. Что касается тех клеток, которые я описывал в этой главе, то очевидно, что они имеют самое прямое отношение к зрительному восприятию. Как я уже упоминал вначале, разрушение любого небольшого участка стриарной коры приводит к слепоте в каком-то небольшом участке поля зрения. У обезьян повреждение стриарной коры дает такой же эффект. Однако у кошек дело обстоит сложнее — кошка с удаленной стриарной корой может видеть, хотя и не так хорошо, как раньше. Другие отделы мозга, такие как верхние бугорки четверохолмия, возможно, играют у нее более важную роль в зрительном восприятии, чем у приматов. У таких низших позвоночных, как лягушки и черепахи, нет ничего похожего на нашу кору большого мозга, однако никто не станет утверждать, что они слепы.
Теперь мы с достаточной уверенностью можем сказать, что именно любая из описанных выше корковых клеток делает в ответ на стимуляцию изображением видимой сцены. Большинство корковых клеток плохо отвечает на диффузное освещение и хорошо — на линии с нужной ориентацией. Так, при показе фигуры, напоминающей по форме почку (рис. 57), такого рода клетка будет отвечать тогда и только тогда, когда участок границы с определенной ориентацией будет пересекать ее рецептивное поле. Те же клетки, рецептивные поля которых расположены внутри границ фигуры, никак не отреагируют — они будут продолжать давать спонтанный импульсный разряд независимо от присутствия или отсутствия данной фигуры.
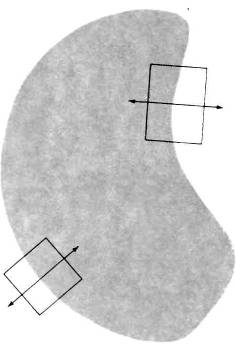
Рис. 57. Насколько вероятно, что клетки в нашем мозгу будут отвечать на те или иные характерные стимулы, такие, например, как изображенная здесь почковидная фигура? Из всего множества клеток зрительной коры реакцию на подобный стимул будет давать только очень небольшая группа клеток.
Так ведут себя клетки с ориентационной избирательностью. Однако для возбуждения простой клетки недостаточно того, чтобы участок контура соответствовал оптимальной ориентации, — контур должен еще почти точно попадать на границу тормозной и возбуждающей зон рецептивного поля, ведь для ответа нужно, чтобы свет падал на возбуждающую зону, но не распространялся на тормозную. Если хотя бы немного сдвинуть участок контура, не меняя его ориентации, стимуляция данной клетки окажется недостаточной, и теперь уже начнет возбуждаться другая популяция простых клеток. Для сложных клеток условия возбуждения не столь жестки, поскольку популяция клеток, активированная стимулом в какой-то момент времени, не изменится при небольшом сдвиге границ фигуры без изменения их ориентации. Чтобы заметно изменить популяцию возбужденных сложных клеток, нужно достаточно сильно сдвинуть границу — так, чтобы она полностью вышла из рецептивных полей одних клеток и попала в поля других. Таким образом, популяция возбужденных сложных клеток — в отличие от простых — в целом мало меняется при небольших поступательных смещениях объекта.
Если, наконец, обратиться к клеткам, реагирующим на концы линий, то и здесь мы обнаружим менее жесткие ограничения на точное положение стимула (хотя каждая данная конфигурация будет активировать гораздо меньше клеток). Для нейронов этого типа ориентация контура должна всегда совпадать с оптимальной ориентацией возбуждающей зоны рецептивного поля, но должна заметно меняться за пределами этой зоны, чтобы торможение не уравновесило возбуждение. Короче говоря, участок контура должен иметь достаточную кривизну или резко прерываться, чтобы были соблюдены все условия для возбуждения данной клетки (см. рис. 56).
Эти жесткие требования повышают избирательность реакций коры, так как каждый видимый объект возбуждает лишь очень малую долю клеток, на рецептивные поля которых падает его изображение. Такая специализация клеток, вероятно, продолжает возрастать и при дальнейшем переходе к еще более высоким уровням за пределами стриарной коры. На палочки и колбочки воздействует просто свет как таковой. Ганглиозные клетки сетчатки, клетки НКТ и корковые нейроны с центром и периферией сравнивают соответствующие участки поля зрения с окружающим фоном, поэтому они, вероятно, будут реагировать на любой отрезок контура, попавшего в их рецептивное поле, но не будет давать ответа на общее изменение освещенности сетчатки. Клетки, избирательно чувствительные к ориентации, отмечают не только наличие контура, но и его ориентацию и даже быстроту ее изменения, т.е. кривизну линии. Если такие клетки относятся к категории сложных, то они чувствительны также к движению. Как говорилось в одном из предыдущих разделов книги, возможны два предположения относительно роли чувствительности к движению: возможно, она позволяет легче обращать внимание на движущиеся объекты, а может быть, этот механизм в сочетании с микросаккадами поддерживает реакцию сложных клеток на неподвижные объекты.
Мне кажется, что границы между светлым и темным — самый важный компонент наших зрительных восприятий, но, без сомнения, далеко не единственный. Цвет различных объектов, безусловно, помогает при выделении их (хотя нужно сказать, что наши последние работы указывают на меньшую роль цвета при определении формы). При зрительной оценке формы и глубины используются также распределение светотени на поверхности предметов и текстура их поверхности. Хотя клетки, работу которых мы обсуждали, в принципе могли бы участвовать в восприятии светотеневых переходов и текстуры, не следует ожидать, что они смогут четко реагировать на оба этих признака. Еще предстоит выяснить, каков механизм восприятия текстур в зрительной системе. Одна из возможных гипотез состоит в том, что сложные клетки обрабатывают информацию о полутонах и текстурах самостоятельно, без помощи каких бы то ни было специализированных групп клеток. Возможно, что такие стимулы не способны сильно активировать большое число клеток, однако текстурам и полутонам свойственна пространственная протяженность, и они могли бы вызывать реакцию сразу многих клеток, хотя каждая клетка будет отвечать умеренно или слабо. Быть может, вялые ответы множества клеток были бы достаточны для передачи информации на более высокие уровни.
Многие люди, в том числе и я, все еще с трудом воспринимают то, что внутренняя часть любой фигуры (например, область внутри почковидного пятна на рис. 57) не является стимулом для клеток нашего мозга и что восприятие однородного внутреннего поля как белого или черного (или окрашенного, как мы увидим в главе 8) зависит только от возбуждения клеток, чувствительных к границам. Логическим доводом в пользу этого служит следующее соображение: если бы при восприятии внутренней области активировались клетки, рецептивные поля которых лежат в ее пределах, то она должна была бы, наоборот, восприниматься как неоднородная! Таким образом, если мы видим все занятое фигурой пространство как равномерно черное, белое, серое или зеленое, то клетки с рецептивными полями внутри границ фигуры не имеют к этому никакого отношения. С этой мыслью трудно смириться, не так ли? Однако с точки зрения инженера, конструирующего машину для кодирования формы, такой механизм, я думаю, был бы очень подходящим. Единственная информация, которую необходимо в этом случае получать, — это информация о внешнем контуре фигуры; анализ ее внутренности становится излишним. Нужно ли удивляться, что эволюция мозга пошла по пути, позволяющему перерабатывать информацию с помощью минимального количества клеток?
Услышав о свойствах простых и сложных клеток, люди часто делают вывод, что для полного анализа всех мельчайших элементов зрительного поля, включая темные и светлые линии и края различных участков во всех возможных ориентациях, требуется астрономическое число клеток. Разумеется, так оно и есть. Но все дело в том, что кора как раз и содержит астрономическое число клеток. Сегодня мы уже можем сказать, как работают клетки в данном участке мозга, по крайней мере как они отвечают на множество простых зрительных стимулов, встречающихся в повседневной жизни. Я подозреваю, что в стриарной коре нет двух клеток, которые работали бы совершенно одинаково. Действительно, всякий раз, когда с помощью микроэлектрода удавалось одновременно регистрировать активность двух клеток, оказывалось, что эти клетки хоть немного, но отличаются друг от друга — по положению рецептивных полей, дирекциональной чувствительности, интенсивности ответов или каким-то другим параметрам. Короче говоря, складывается впечатление, что в данном отделе мозга если и есть избыточность, то она невелика.
Можем ли мы быть уверены в том, что описанные клетки на самом деле предназначены именно для выявления отрезков прямых линий, а не каких-то других стимулов? Нельзя сказать, что мы и другие исследователи не пытались применять множество других стимулов, включая лица, карту мира и пассы руками. Как показывает опыт, было бы глупо думать, что мы исчерпали все возможности. В начале 1960-х годов, когда мы удовлетворились результатами, полученными с клетками стриарной коры, и решили перейти (и даже фактически уже перешли) к следующей области, нам случайно удалось записать реакцию одной слабо отвечавшей клетки стриарной коры. Однако, сделав светлую линию более короткой, мы убедились, что эта клетка способна давать весьма энергичную реакцию. Именно тогда мы наткнулись на класс клеток, отвечающих на концы линий. После этого мы еще почти двадцать лет работали с корковыми клетками обезьян, прежде чем обнаружили «пузырьки» — скопления клеток, специфически реагирующих на цвет (они описаны в главе 8). Сделав эти оговорки, я должен добавить, что некоторые из свойств, обнаруженных у клеток стриарной зоны, например ориентационная чувствительность, являются, несомненно, истинными свойствами этих клеток. В пользу этого можно привести много дополнительных данных, например данные по функциональной анатомии, описанные в главе 5.
Бинокулярная конвергенция
До сих пор я почти не упоминал о существовании двух глаз. Очевидный интерес представляет вопрос: получает ли та или иная корковая клетка входные сигналы от обоих глаз, и если да, то одинаковы ли эти входы в количественном и качественном отношении?
Для того чтобы получить ответ, мы должны вернуться на некоторое время к наружному коленчатому телу (НКТ) и выяснить, имеют ли какие-то из его клеток входы от обоих глаз. НКТ — это самый низкий уровень, на котором уже возможно было бы объединение сигналов от двух глаз на одной клетке. Однако возможность эта, по-видимому, здесь не реализована — два разных пучка входных волокон распределяются по разным клеточным слоям, между которыми нет или почти нет взаимодействия. Как можно было бы ожидать, учитывая раздельность проекций от двух глаз, отдельная клетка НКТ должна отвечать на стимуляцию одного какого-то глаза и совсем не реагировать на стимуляцию другого. Судя по результатам некоторых экспериментов, стимулы, подаваемые на «чужой» для данной клетки НКТ глаз, могут оказывать слабое воздействие на реакцию, вызываемую со «своего» глаза. Однако практически можно считать, что каждая клетка находится под контролем только одного глаза.
Интуитивно ясно, что пути, идущие от каждого глаза, должны рано или поздно сойтись вместе, так как мы, глядя на что-либо, видим одну целостную картину. Тем не менее повседневный опыт говорит нам, что если закрыть один глаз, то большого изменения не произойдет — предметы будут казаться такими же четкими, такими же реальными и такими же яркими. Разумеется, общее поле зрения для двух глаз будет несколько шире, поскольку каждый глаз видит на своей стороне бо?льшую область пространства, чем другой глаз, хотя эта разница составляет всего лишь около 20–30°. Существенное отличие бинокулярного зрения от монокулярного состоит в ощущении глубины, о чем пойдет речь в главе 7.
В коре обезьяны те клетки, которые получают входные сигналы от НКТ и имеют рецептивные поля с круговой симметрией, сходны с клетками НКТ в том, что они тоже монокулярные. На этом корковом уровне мы находим примерно одинаковое число клеток, возбуждающихся от левого и от правого глаза, по крайней мере в тех участках коры, которые обслуживают область поля зрения в радиусе около 20° от центральной точки фиксации. Однако на следующем уровне коры выявляются уже бинокулярные клетки, простые и сложные, причем у макаков больше половины таких клеток могут реагировать независимо на сигналы от каждого из глаз.
Как только мы нашли бинокулярную клетку, можно тщательно сравнить ее рецептивные поля в обеих сетчатках. Сначала закрываем правый глаз животного и картируем рецептивное поле клетки в левом глазу, отмечая его точное положение на экране или на сетчатке, а также его сложность, ориентацию и расположение возбуждающих и тормозных зон; выясняем также, простая это клетка или сложная, исследуем ее способность реагировать на конец линии и дирекциональную чувствительность; после этого закрываем левый глаз животного, открываем правый и повторяем всю процедуру измерений сначала. Оказалось, что у большинства бинокулярных клеток все свойства, выявляемые в опытах с левым глазом, обнаруживаются и при стимуляции правого глаза — то же положение на сетчатке, та же дирекциональная чувствительность и т.д. Это позволяет заключить, что все связи, идущие к данной клетке от левого глаза, совпадают по структуре со связями, идущими от правого глаза.
Говоря о таком дублировании связей, нужно сделать одно уточнение. Если, определив для клетки оптимальный стимул, его положение, ориентацию, направление движения и т.д., мы сравним ее ответы при стимуляции одного глаза и при стимуляции другого, интенсивность реакции не всегда окажется одинаковой. Некоторые клетки действительно одинаково хорошо активируются от обоих глаз, однако другие явно дают более сильный разряд при стимуляции определенного глаза. В целом, за исключением той части корковых клеток, которые обслуживают периферию поля зрения, мы не находим никакого особого преимущества того или другого глаза — в каждом полушарии число клеток, лучше активируемых с противоположной стороны (от контралатерального глаза) и с той же стороны (от ипсилатерального глаза), примерно одинаково. При этом встречаются все степени относительного доминирования глаза, начиная от клеток, возбуждаемых исключительно с левого глаза, и кончая клетками, отвечающими только на стимуляцию правого глаза.
Теперь можно оценить численность различных популяций клеток. Разобьем все изученные клетки, скажем 1000 штук, произвольно на семь классов по относительной эффективности воздействия на них того или другого глаза. Затем подсчитаем число клеток в каждом классе. На рис. 58 представлены соответствующие гистограммы для кошки и макака. Здесь сразу видны сходства и различия в распределении клеток у этих животных. Видно, что у обоих видов бинокулярные клетки встречаются достаточно часто, причем среди клеток с односторонним доминированием хорошо представлены оба глаза (у макака примерно поровну), что у кошек бинокулярных клеток очень много, что у макаков численность монокулярных и бинокулярных клеток примерно одинакова, причем у бинокулярных клеток зачастую сильно выражено доминирование одного глаза (группы 2 и 5), и что реже всего встречаются клетки, одинаково хорошо активируемые с любого глаза.
Рис. 58. Изучая распределение нейронов по глазодоминантности, мы исследовали сотни клеток и относили каждую из них к одной из семи произвольно выделенных групп. Клетки группы 1 определяются как клетки, на которые влияет только контралатеральный глаз, т.е. глаз, находящийся на противоположной стороне тела; клетки группы 2 реагируют на стимуляцию обоих глаз, однако явно предпочитают сигналы от контралатерального глаза; и т.д.
Рис. 59. Регистрирующий электрод оказался достаточно близко к трем клеткам, чтобы отводить импульсы от всех трех. Ответы разных клеток можно различить по амплитуде и форме импульсов. На рисунке показаны реакции на стимулы, предъявленные одному глазу и обоим глазам. Клетки 1 и 2 должны быть отнесены к группе 4, так как они почти одинаково отвечают на стимуляцию того и другого глаза. Клетка 3 отвечает только на одновременную стимуляцию обоих глаз; можно сказать лишь то, что она не принадлежит ни к группе 1, ни к группе 7.
Теперь мы можем продвинуться дальше и задать следующий вопрос: отвечают ли бинокулярные клетки при стимуляции обоих глаз лучше, чем при стимуляции одного глаза? Оказывается, многие клетки именно таковы — от одного глаза они активируются слабо или не возбуждаются совсем, а от двух глаз дают сильный импульсный разряд, особенно в том случае, когда оба глаза стимулируются одновременно и совершенно одинаково. На рис. 59 показаны записи ответов трех клеток (1, 2 и 3), демонстрирующих выраженную синергию. Одна из этих клеток вообще не отвечает на стимуляцию только одного глаза, так что мы вообще не смогли бы ее обнаружить, если бы не стимулировали оба глаза одновременно. Эффект синергии у многих клеток выражен слабо или не отмечается вовсе — такие клетки отвечают на стимуляцию обоих глаз примерно так же, как и на стимуляцию каждого глаза по отдельности.
Подобные связи одиночных клеток с двумя глазами еще раз указывают на высокую степень специфичности соединений в мозгу. Мало того, что входные системы связей данной клетки позволяют ей отвечать только на линию определенной ориентации и лишь на одно направление движения, — оказывается к тому же, что эти системы представлены двумя копиями, по одной от каждого глаза. Но и этого еще мало: как мы узнаем из главы 9, большинство связей, по-видимому, должно быть сформировано и готово к работе уже к моменту рождения животного. Все это поистине удивительно.
Глаз, мозг, зрение
Дэвид Хантер Хьюбел; David Hunter Hubel;
Дек 24 2017