
Статьи. Предметно
«Увеличение» и модули
«Увеличение» и модули
В предыдущей главе я подчеркивал, что первичная зрительная кора кажется морфологически однородной как невооруженному глазу, так и под микроскопом при использовании большинства обычных методов окрашивания. Однако при более тщательном изучении выяснилось, что однородна и топография колонок глазодоминантности: период чередования зон доминирования левого и правого глаза остается на удивление постоянным от проекции центральной ямки (точки фиксации) до дальней периферии бинокулярной части поля зрения. При помощи метода с инъекцией дезоксиглюкозы мы выявили также однородность топографии ориентационных колонок.
Факт анатомической однородности коры мы вначале восприняли как неожиданность, так как с функциональной точки зрения эта область явно неоднородна по двум важным параметрам. Во-первых, как уже говорилось в главе 3, рецептивные поля ганглиозных клеток сетчатки в зоне центральной ямки и около нее гораздо меньше, чем на периферии. Размеры рецептивного поля типичной сложной клетки в верхнем слое коры в области проекции центральной ямки составляют примерно 0,25?0,5°. Если же рассматривать те части коры, где представлена периферия сетчатки (80–90° от центральной ямки), то здесь чаще всего встречаются рецептивные поля размером 2?4°. Если сравнивать соответствующие площади, то отношение между ними можно оценить как 1:10 или даже 1:30.
Второй вид неоднородности связан с так называемым «увеличением». Это понятие, введенное в 1961 году П. Дэниелом и Д. Уиттериджем, означает расстояние между двумя точками коры, в которые проецируются две точки поля зрения, разделенные расстоянием в 1°. Если идти от центральной ямки к периферии, корковая проекция одного и того же углового расстояния будет становиться все меньше и меньше, т.е. «увеличение» будет уменьшаться. Если сместить стимул на 1° около точки фиксации, то его проекция в коре сдвинется примерно на 6 мм. Если же поместить этот стимул в зону, отстоящую от точки фиксации на 90°, то сдвиг в поле зрения на 1° будет соответствовать сдвигу вдоль поверхности коры всего лишь на 0,15 мм. Таким образом, «увеличение» в центре сетчатки примерно в 36 раз больше, чем на периферии.
Обе рассмотренные неоднородности связаны с одним и тем же обстоятельством — с уменьшением остроты зрения по мере удаления от центра сетчатки к ее периферии. Попробуйте, например, фиксировать взором какую-нибудь букву в самом начале строки и попытайтесь определить, какая буква или слово находится в самом ее конце. Или фиксируйте взглядом букву п в начале слова прогрессивно. Вряд ли вам удастся опознать букву о в конце слова. Вероятно, вам трудно будет опознать также в и н перед последней буквой. Для того чтобы обеспечить высокое разрешение в центральной ямке, зрительной системе требуется много корковых клеток на единицу площади в проекции этого участка, причем нужно, чтобы каждая такая клетка имела очень небольшое рецептивное поле.
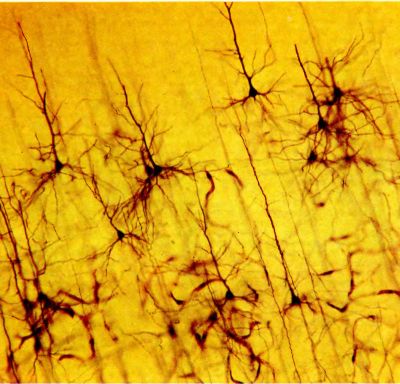
Рис. 90. В этой главе речь пойдет о «модулях». Один модуль по размерам примерно соответствует изображенному здесь срезу зрительной коры, окрашенному по Гольджи. Этот метод окраски выявляет лишь очень малую долю всех нервных клеток, но если уж клетка оказывается окрашенной, то она видна вся полностью или почти вся: на таком срезе можно рассмотреть тело клетки, ее дендриты и аксон.
Разброс и смещение рецептивных полей
Учитывая сказанное выше, как можно объяснить анатомическую однородность коры? Для этого придется подробнее рассмотреть, что? происходит с положением рецептивных полей нейронов, встречающихся на пути продвижения электрода. Если мы будем вводить электрод в стриарную кору точно под прямым углом к ее поверхности, то окажется, что все эти рецептивные поля расположены почти в одном и том же участке поля зрения, но не совпадают полностью — от клетки к клетке положение рецептивного поля несколько меняется. Однако эти изменения кажутся случайными и не очень велики, так что рецептивные поля каждых двух последовательных клеток значительно перекрываются (этот факт отражен на рис. 91). В каждом данном слое коры размеры рецептивных полей относительно постоянны, однако при переходе к другому слою они заметно меняются — от очень малых в слое 4C до больших в слоях 5 и 6. В пределах одного слоя коры площадь, занимаемая десятью или двенадцатью последовательно зарегистрированными полями, из-за случайного разброса будет примерно в 2–4 раза больше, чем площадь рецептивного поля одной клетки. Назовем ту область, которую занимает вся совокупность рецептивных полей отдельных клеток в одном слое и в одной точке коры составным рецептивным полем данной точки. В зависимости от слоя коры величина составного поля варьирует — например, в слое 3 она равна около 30 угловых минут для центральной ямки и примерно 7–8° для дальней периферии.
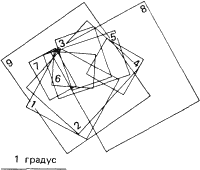
Рис. 91. Картирование этих девяти рецептивных полей было проведено при однократном прохождении микроэлектрода через стриарную кору кошки перпендикулярно к ее поверхности. При таком продвижении электрода наблюдаются случайные отклонения в положении рецептивных полей, а также в их величине, но не отмечается какой-либо тенденции к их систематическому смещению.
Теперь предположим, что мы вводим и продвигаем электрод горизонтально в пределах одного слоя коры, скажем слоя 3. И здесь при последовательной регистрации ответов клеток мы обнаружим, что положение рецептивных полей варьирует случайным образом, однако в этих беспорядочных сдвигах мы заметим тенденцию к смещению в определенном направлении. Разумеется, это направление можно предсказать исходя из топографической проекции поля зрения в кору. Сейчас нас будет интересовать величина смещения, которая наблюдается при сдвиге электрода на 1 миллиметр по горизонтали. Если учесть то, что было сказано выше относительно различий в «увеличении», ясно, что расстояние, измеряемое в поле зрения, будет зависеть от того, где именно в коре мы продвигаем электрод — в области проекции центральной ямки, в периферийной области или где-то в промежуточной зоне. Скорость смещения рецептивных полей в поле зрения будет далеко не одинаковой. Оказывается, однако, что эта скорость будет очень близка к постоянной, если сделать поправку на размер самих рецептивных полей. Смещение в любом месте коры на 1 миллиметр соответствует такому сдвигу в поле зрения, который равен примерно половине поперечника составного рецептивного поля (т.е. суммарного поля, образуемого наложением рецептивных полей всех клеток, находящихся под данной точкой поверхности коры. Таким образом, чтобы полностью выйти из одного участка поля зрения и перейти в другой, следующий, потребуется смещение примерно в 2 миллиметра (это показано на рис. 92). То же оказывается справедливым для всех точек поля 17, где мы отводили электрическую активность. В области центральной ямки рецептивные поля очень малы, однако невелик и сдвиг в поле зрения, который соответствует смещению вдоль коры на 2 миллиметра. Напротив, на периферии как размеры рецептивных полей, так и соответствующие смещения в поле зрения гораздо больше (рис. 93).
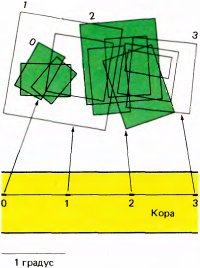
Рис. 92. При продвижении электрода в коре кошки на большое расстояние параллельно поверхности положение рецептивных полей встречающихся клеток изменяется. В данном случае электрод прошел 3 мм и встретился примерно с 60 клетками (их слишком много, чтобы показать на этом рисунке). Здесь отмечены положения только четырех или пяти рецептивных полей, карты для которых составлялись в начале прохождения каждого миллиметра (примерно десятой его части), а остальные поля оставлялись без внимания. В верхней части рисунка показаны рецептивные поля клеток, встретившихся на четырех этапах проходки (0, 1, 2 и 3). Каждая такая группа полей заметно смещалась в поле зрения вправо относительно предыдущей группы. Рецептивные поля клеток группы 2 не перекрываются с полями группы 0, а поля группы 3 — с полями группы 1. В обоих случаях расстояние вдоль коры составляло 2 миллиметра.
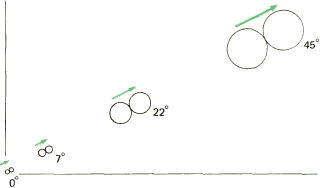
Рис. 93. У макака рецептивные поля клеток верхних слоев коры увеличиваются по мере удаления от центральной ямки (которой соответствует отметка 0°). В таком же отношении увеличивается и расстояние между центрами рецептивных полей тех клеток, расстояние между которыми в коре составляет 2 миллиметра.
Функциональные единицы коры
Нам приходится заключить, что любой участок первичной зрительной коры величиной примерно 2?2 миллиметра должен иметь все механизмы, необходимые для полной обработки определенного участка поля зрения, который очень мал в области центральной ямки и гораздо больше на периферии. Такой участок, получающий, вероятно, десятки тысяч входных волокон от наружного коленчатого тела, перерабатывает поступающую по ним информацию и посылает выходные сигналы по волокнам, чувствительным к ориентации стимула, его движению и т.д., объединяя информацию от обоих глаз. Все участки производят сходные операции над сигналами, поступающими примерно по одному и тому же числу волокон. Информация, получаемая отдельным участком, весьма детальна, если поступает с малой площади в области центральной ямки, и менее подробна, когда приходит с большей площади с периферии сетчатки. Выходные сигналы такого участка вырабатываются без всякого учета того, какой по величине участок поля зрения здесь анализируется и с какой степенью детализации. Набор механизмов для переработки информации в таких участках в основном один и тот же. Это позволяет объяснить ту однородность, которую мы видим и невооруженным глазом, и при микроскопическом исследовании.
Тот факт, что сдвига вдоль коры на 2 мм как раз достаточно, чтобы попасть на проекцию нового участка сетчатки, означает, что какие бы локальные операции ни выполнялись в коре, все они должны выполняться в пределах блока размером 2?2 мм. Очевидно, меньший участок не сможет всесторонне анализировать соответственно меньшую площадку поля зрения, так как для такого анализа нужны все элементы 2-миллиметрового блока. Это ясно уже из рассмотрения данных о положении и размерах рецептивных полей. Однако нам надо будет подробнее обсудить, что означают слова «анализ» и «операция». Можно начать с рассмотрения параметра ориентации линии. В любом участке поля зрения, даже самом малом, должны быть учтены все возможные ориентации. Если при анализе какого-то участка соответствующим корковым блоком величиной 2 мм в этом блоке не окажется элемента, реагирующего на ориентацию +45°, то наличие такого элемента в других блоках коры не поможет, так как другие блоки имеют дело с другими участками поля зрения. Однако, к счастью, ширина ориентационных полос в коре достаточно мала — 0,05 мм, и набор таких полос для всех ориентаций от 0 до 180° с шагом в 10° дважды укладывается на участке коры шириной 2 мм. Так же обстоит дело и с глазодоминантностью — ширина соответствующих колонок 0,5 мм, поэтому 2-миллиметрового блока более чем достаточно для полного анализа. Таким образом, блоки поперечником в 2 мм обладают, по-видимому, полным набором необходимых механизмов.
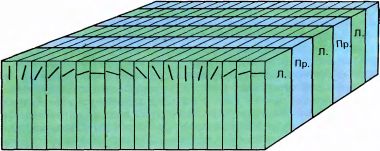
Рис. 94. Мы называем эту схему «нашей моделью из ледяных кубиков». Схема показывает, как разбита кора одновременно на два вида пластин — для глазодоминантности (для правого и левого глаза) и для ориентации. Эту модель не следует воспринимать буквально — ни одна из этих систем пластин не бывает такой регулярной, особенно ориентационные пластины, которые далеки от того, чтобы располагаться параллельно и иметь прямые границы. Более того, они, по-видимому, не пересекаются под каким-то определенным углом и уж во всяком случае не образуют ортогональную систему вроде показанной на рисунке.
Следует добавить, что блоки величиной 2 мм — это особенность не столько всего поля 17 коры, сколько слоя 3 поля 17. Если обратиться к слоям 5 и 6, то там размеры рецептивных полей клеток и их разброс вдвое больше; поэтому для всестороннего анализа в слоях 5 и 6 и образования рецептивных полей большей величины и с более сложными свойствами, по-видимому, потребуются блоки с поперечником в 4 мм. С другой стороны, в слое 4C рецептивные поля и их разброс гораздо меньше и соответственно размеры блоков ближе к 0,1–0,2 мм. Однако общая идея отдельных функциональных блоков остается прежней независимо от того факта, что любая заданная область поля зрения обслуживается набором локальных механизмов, размещенных в нескольких слоях коры, т.е. что кора — это совокупность нескольких систем, объединенных в одно целое.
Все эти соображения помогают нам понять, почему колонки не могут быть гораздо более крупными. В блок размером 2?2 мм нужно поместить все механизмы, выделяющие нужные переменные в стимулах. До сих пор мы говорили об ориентации и глазодоминантности, однако на самом деле на двумерную поверхность отображаются не две, а большее число входных переменных. Для этого в качестве основных параметров выбраны две переменные, определяющие координаты в поле зрения (расстояния от центральной ямки по горизонтали и по вертикали). На полученной координатной сетке в виде более тонкой мозаики дополнительно отображены и другие переменные, такие как ориентация стимула и глазодоминантность.
Мы будем называть участок коры с поперечником 2 мм модулем. Лично мне этот термин кажется не вполне подходящим, в частности, потому, что он вызывает слишком конкретное представление о небольшом прямоугольном блоке электронного устройства, который вместе с сотнями других таких же блоков смонтирован на общем шасси. Однако в определенной мере мы имели в виду именно эту ассоциацию, но более обобщенного характера. Во-первых, выделенные нами функциональные модули ориентационной системы таковы, что границы между смежными модулями условны. Например, можно считать, что модуль начинается с клеток, выделяющих вертикальную ориентацию, и заканчивается следующей группой клеток, которые тоже выделяют вертикальную ориентацию; за начало можно принять и любую другую ориентацию, лишь бы в каждом модуле все ориентации были представлены хотя бы по одному разу. Так же обстоит дело и с глазодоминантностью — модуль может начинаться с того места, где доминирует правый глаз, с того, где доминирует левый, или с середины колонки, лишь бы в конечном счете в модуль вошли два участка глазодоминантности, по одному для каждого глаза. Во-вторых, как мы уже упоминали, величина модуля будет зависеть от того, какой из слоев коры мы рассматриваем. Тем не менее термин «модуль» должен ассоциироваться с системой примерно из 500–1000 миниатюрных устройств, которые все взаимозаменяемы (при условии, что можно было бы воспроизвести около 10 000 входных и, вероятно, около 50 000 выходных связей).
Необходимо тут же добавить, что никто, конечно, не думает, что кора совершенно однородна, — от проекции центральной ямки до участков, отображающих дальнюю периферию. Помимо остроты зрения по мере удаления от центра сетчатки изменяются и другие параметры зрительного восприятия; ухудшается, например, цветовое зрение, хотя, возможно, и не очень резко, если компенсировать уменьшение коэффициента «усиления» (т.е. увеличивать размеры объектов по мере удаления их от точки фиксации). Движение, равно как и очень слабые источники света, по всей вероятности, легче замечается на периферии поля зрения. Бинокулярное зрение должно ухудшаться, так как в зонах радиусом от 20 до 80° колонки глазодоминантности для ипсилатерального глаза становятся все уже и уже, а для контралатерального глаза — все шире и шире. На расстояниях больше 80° от центральной ямки ипсилатеральные колонки полностью исчезают и корковые механизмы становятся «монокулярными». Ввиду этих, а также и других различий, которые, несомненно, имеются, соответствующие корковые структуры должны быть в чем-то неоднородными. Поэтому корковые модули, вероятно, не все совершенно одинаковы.
Деформация коры
Если сравнивать кору с сетчаткой, то можно получить более полное представление о конфигурации коры. Глаз представляет собой сферу, поскольку это чисто оптический инструмент. Поэтому форма сетчатки тоже сферическая. В фотоаппарате пленка может быть плоской, так как апертура линзы составляет в среднем около 30°. В широкоугольной камере типа «рыбий глаз» апертура гораздо больше, однако на периферии изображение искажается. Конечно, фотографии сферической формы были бы неудобны — плоские фотоснимки гораздо удобнее хранить. Что касается глаза, то его шаровидная форма представляется идеальной, так как шар — тело компактное, и в то же время он может легко вращаться в глазнице (если бы глаз имел форму куба, то возникли бы затруднения). А поскольку глаз имеет сферическую форму, то угловой масштаб изображения зрительного поля на сетчатке остается везде постоянным — число градусов, приходящееся на 1 мм сетчатки, на всей сетчатке одинаково. У человека оно составляет 3,5°. Как уже говорилось, центры рецептивных полей ганглиозных клеток в центральной ямке и поблизости от нее очень малы, но они увеличиваются по мере удаления от нее. Поэтому мы не должны удивляться тому, что в центральной области сетчатки в каждом миллиметре должно быть гораздо больше ганглиозных клеток, чем в зоне дальней периферии. Действительно, вблизи центральной ямки эти клетки уложены несколькими слоями, тогда как на периферии сетчатки располагаются настолько редко, что визуально не образуют даже одного непрерывного слоя (см. микрофотографии на рис. 95). Поскольку сетчатка должна иметь сферическую форму, то ее слои не могут быть однородными. Возможно, это одна из причин того, что в сетчатке происходит лишь весьма ограниченная обработка информации; иначе потребовалось бы очень большое утолщение ее слоев в центральном участке.
В этом смысле кора обладает бо?льшими возможностями. В отличие от сетчатки она может не быть сферической, а может просто растягиваться в области проекции центральной ямки по сравнению с проекцией периферии. Это растяжение, по-видимому, осуществляется в такой степени, чтобы толщина коркового слоя (так же как ширина колонок и другие параметры) была повсюду одинаковой.
Какое влияние это оказывает на общую форму коры? Хотя я неоднократно характеризовал кору как «слой», это не означает, что она должна быть плоской. Если бы не было никаких искажений конфигурации, стриарная кора имела бы сферическую форму — такую же как у глазного яблока. В этом смысле она похожа на поверхность Земли — без искажений, связанных с рельефом, поверхность Земли имела бы строго сферическую форму. Правда, на стриарную кору одного полушария отображается лишь около половины задней поверхности глаза, т.е. примерно четверть всей сферы.
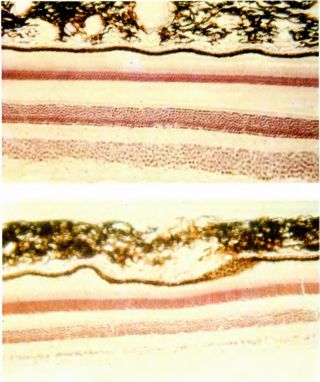
Рис. 95. В отличие от слоев коры слои сетчатки далеко не одинаковы по толщине в различных ее участках. Как у обезьяны, так и у человека слой ганглиозных клеток вблизи центральной ямки (нижний слой на верхней микрофотографии) имеет толщину в 8–10 клеток, в то время как на дальней периферии сетчатки, на расстоянии 70–80° от ее центра (нижняя микрофотография) ганглиозные клетки не образуют даже одного сплошного слоя. Это не должно нас удивлять, поскольку центры рецептивных полей ганглиозных клеток около центральной ямки очень малы, а на периферии они увеличиваются. В центре сетчатки на единицу площади приходится гораздо больше ганглиозных клеток.
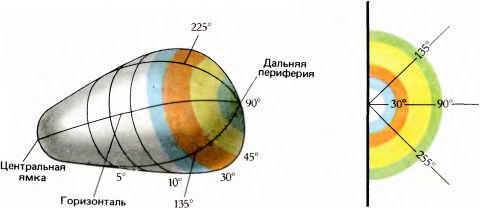
Рис. 96. Расправленная стриарная кора по форме напоминала бы грушу. Она имела бы вид четверти сферы, если бы все части сетчатки были представлены в ней пропорционально. На самом деле, однако, форма коры значительно искажена из-за несоразмерно увеличенного отображения центральной области сетчатки. Дальняя периферия (примерно в 90° от центра) отображена так слабо, что полукруг, соответствующий 90° (крайний справа), оказывается очень небольшим.
Растягиваясь таким образом, чтобы сохранять постоянную толщину и все же иметь возможность переработать особенно обильную информацию от множества ганглиозных клеток центральной ямки, кора приобретает форму, отличную от сферической. Если развернуть и разгладить все складки коры, форма ее не окажется ни сферической, ни плоской. В действительности кора будет тогда иметь вид сильно искаженного сегмента сферы, составляющего четвертую ее часть; по форме она скорее будет напоминать грушу или яйцо. Такой результат предсказывали в 1962 году Дэниел и Уиттеридж, экспериментально определявшие зависимость «увеличения» в поле 17 от расстояния до проекции центральной ямки (об этом говорилось на с. 0127). Из полученных данных они вычислили трехмерную форму поверхности коры, а затем по серии последовательных гистологических срезов сделали резиновую модель. После этого они на самом деле расправили эту модель, тем самым подтвердив свое предсказание относительно грушевидной формы коры. Полученная форма показана на рис. 96. До этого никто не задавался вопросом, какую форму будет иметь зрительная кора, если расправить ее складки. Насколько мне известно, никто также не сознавал, что любая область коры должна иметь какую-то определенную форму, логически вытекающую из функций этой области. По-видимому, складки коры, которые для выяснения истинной формы нужно разгладить (без растяжения и разрывов), образуются из-за того, что большой сегмент коры должен был быть «скомкан», чтобы поместиться в ограниченном пространстве черепной коробки. При этом характер складок может быть не совсем случайным; какие-то детали, вероятно, определяются тем, что межкортикальные связи должны быть по возможности короче.
Если обратиться к соматосенсорной коре, то здесь сложность топографического отображения может доходить до крайней степени. Например, область коры, соответствующая коже руки, по конфигурации должна быть похожа на перчатку, причем с определенными искажениями, чтобы обеспечить бо?льшую чувствительность кончиков пальцев по сравнению с ладонью и тыльной стороной кисти. Такое искажение аналогично увеличению корковой проекции центральной ямки по сравнению с периферией сетчатки в соответствии с различием в остроте зрения. Будет ли корковая проекция руки действительно напоминать перчатку, если мы сделаем резиновую модель и будем ее постепенно раздувать, чтобы расправить искусственные складки? Вероятно, нет. Картирование соматосенсорной коры оказалось чрезвычайно сложной задачей. Результаты, полученные до сих пор, позволяют думать, что форма непрерывной проекции была бы слишком причудливой; на самом деле соматосенсорная кора как будто ножницами разрезана на множество небольших, более «удобных» участков, которые соединены наподобие кусков лоскутного одеяла так, что получается некоторое приближение к плоской поверхности.
Глаз, мозг, зрение
Дэвид Хантер Хьюбел; David Hunter Hubel;
Дек 24 2017