
Статьи. Предметно
Мозолистое тело и стереоскопическое зрение
Мозолистое тело и стереоскопическое зрение
Мозолистое тело представляет собой мощный пучок миелинизированных волокон, соединяющих два полушария мозга. Стереоскопическое зрение (стереопсис) — это способность воспринимать глубину пространства и оценивать удаленность предметов от глаз. Эти две вещи не особенно тесно связаны друг с другом, однако известно, что небольшая часть волокон мозолистого тела все же играет некоторую роль в стереопсисе. Оказалось удобным включить обе эти темы в одну главу, так как при их рассмотрении придется учитывать одну и ту же особенность устройства зрительной системы, а именно то, что в хиазме имеются как перекрещенные, так и неперекрещенные волокна зрительного нерва (см. рис. 35).
Мозолистое тело
Мозолистое тело (по-латыни corpus callosum) — это самый крупный пучок нервных волокон во всей нервной системе. По приближенной оценке в нем насчитывается около 200 млн. аксонов. Истинное число волокон, вероятно, еще больше, так как приведенная оценка основана на данных обычной световой, а не электронной микроскопии. Это число несравнимо с числом волокон в каждом зрительном нерве (1,5 млн.) и в слуховом нерве (32 000). Площадь поперечного сечения мозолистого тела составляет около 700 мм2, тогда как у зрительного нерва она не превышает нескольких квадратных миллиметров. Мозолистое тело вместе с тонким пучком волокон, называемым передней комиссурой, соединяет два полушария мозга (рис. 98 и 99). Термин комиссура означает совокупность волокон, соединяющих две гомологичные нервные структуры, расположенные в левой и правой половинах головного или спинного мозга. Мозолистое тело тоже иногда называют большой комиссурой мозга.

Рис. 97. На этой стереопаре изображена галерея в Новом колледже в Оксфорде. После того как была сделана сдвинули на 7,5 см влево и сделали левую фотографию.
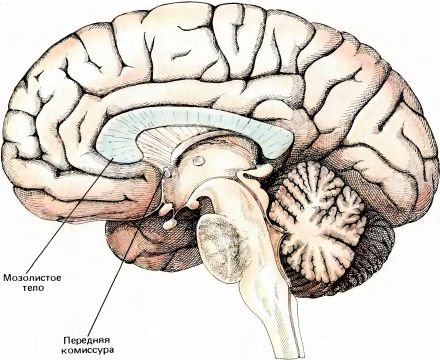
Рис. 98. Головной мозг человека в разрезе, проходящем по срединной плоскости. Виден толстый, дугообразный в поперечном сечении пучок аксонов — мозолистое тело.
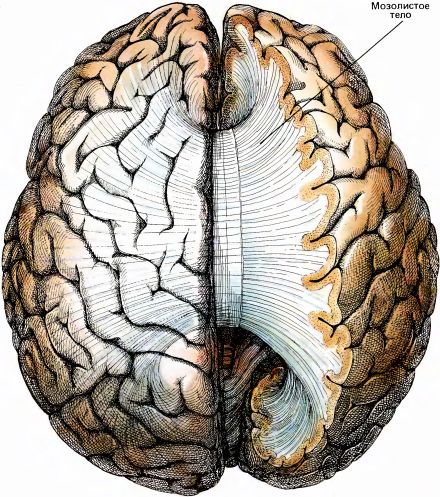
Рис. 99. Здесь мозг показан сверху. Часть правого полушария срезана, и виден пучок волокон мозолистого тела, соединяющий все участки двух полушарий.
Примерно до 1950 года роль мозолистого тела была совершенно неизвестна. В редких случаях наблюдается врожденное отсутствие (аплазия) мозолистого тела. Это образование может также быть частично или полностью перерезано во время нейрохирургической операции, что делается намеренно — в одних случаях при лечении эпилепсии (чтобы судорожный разряд, возникающий в одном полушарии мозга, не мог распространиться на другое полушарие), в других случаях для того, чтобы добраться сверху до глубоко расположенной опухоли (если, например, опухоль находится в гипофизе). По наблюдениям невропатологов и психиатров, после такого рода операций не возникает никаких расстройств психики. Кто-то даже высказал мысль (хотя вряд ли всерьез), что единственная функция мозолистого тела состоит в том, чтобы удерживать два полушария мозга вместе. Вплоть до 1950-х годов мало что было известно о деталях распределения связей в мозолистом теле. Очевидно было, что мозолистое тело соединяет два полушария, и на основании данных, полученных довольно грубыми нейрофизиологическими методами, считали, что в стриарной коре волокна мозолистого тела связывают в точности симметричные участки двух полушарий.
В 1955 году Рональд Майерс, аспирант психолога Роджера Сперри из Чикагского университета, впервые провел эксперимент, в котором удалось выявить некоторые функции этого огромного волокнистого тракта. Майерс занимался обучением кошек, помещенных в ящик с двумя поставленными рядом экранами, на которые можно было проецировать различные изображения, например круг на один экран и квадрат — на другой. Кошку обучали упираться носом в тот экран, на котором было изображение круга, и игнорировать другой — с изображением квадрата. Правильные ответы подкреплялись пищей, а за ошибочные ответы кошек слегка наказывали — включался громкий звонок, и кошку не грубо, но решительно оттаскивали от экрана. Таким методом за несколько тысяч повторений кошку удается довести до уровня надежного различения фигур. (Кошки обучаются медленно; например, голубям для обучения в аналогичной задаче требуется от нескольких десятков до нескольких сотен повторений, а человека вообще можно научить сразу, дав ему словесную инструкцию. Такая разница кажется несколько странной — ведь у кошки головной мозг во много раз больше, чем у голубя.)
Нет ничего удивительного в том, что кошки Майерса научались ничем не хуже решать эту задачу и в том случае, когда один глаз животного был закрыт маской. Неудивительно и то, что если обучение такой задаче, как выбор треугольника или квадрата, проводилось лишь с одним открытым глазом — левым, а при проверке левый глаз закрывали и открывали правый, то точность различения оставалась прежней. Нас это не удивляет потому, что мы сами легко можем решить аналогичную задачу. Легкость решения подобных задач понятна, если учесть анатомию зрительной системы. Каждое полушарие получает входные сигналы от обоих глаз. Как мы уже говорили в главе 4, большая часть клеток в поле 17 тоже имеет входы от обоих глаз. Майерс создал более интересную ситуацию, произведя продольную перерезку хиазмы по средней линии. Таким образом, он перерезал перекрещивающиеся волокна и сохранил в целости неперекрещивающиеся (эта операция требует от хирурга определенного навыка). В результате такой перерезки левый глаз животного оказался соединен только с левым полушарием, а правый — только с правым. Идея эксперимента заключалась в том, чтобы обучать кошку, используя левый глаз, а на «экзамене» адресовать стимул правому глазу. Если кошка сможет правильно решать задачу, то это будет означать, что необходимая информация передается из левого полушария в правое по единственному известному пути — через мозолистое тело. Итак, Майерс произвел продольную перерезку хиазмы, обучил кошку с одним открытым глазом, а затем устроил проверку, открыв другой глаз и закрыв первый. В этих условиях кошки по-прежнему успешно решали задачу. Наконец, Майерс повторил эксперимент на животных, у которых предварительно были перерезаны и хиазма, и мозолистое тело. На этот раз кошки задачу не решили. Таким образом, Майерс опытным путем установил, что мозолистое тело действительно выполняет какие-то функции (хотя вряд ли можно было думать, что оно существует только для того, чтобы отдельные люди или животные с перерезанной зрительной хиазмой могли решать определенные задачи с использованием одного глаза после обучения с использованием другого).
Изучение физиологии мозолистого тела
Одно из первых нейрофизиологических исследований в этой области было проведено спустя несколько лет после экспериментов Майерса Д. Уиттериджем, работавшим тогда в Эдинбурге. Уиттеридж рассудил, что нет особого смысла в том, чтобы пучки нервных волокон соединяли гомологичные зеркально-симметричные участки полей 17. Действительно, не видно никаких причин для того, чтобы нервная клетка в левом полушарии, связанная с какими-то точками в правой половине поля зрения, соединялась с клеткой в правом полушарии, связанной с симметричным участком левой половины поля зрения. Для проверки своих предположений Уиттеридж перере?зал зрительный тракт на правой стороне мозга позади хиазмы и тем самым перекрыл входным сигналам путь в правую затылочную долю; но это, конечно, не исключало передачу туда сигналов из левой затылочной доли через мозолистое тело (рис. 100). Затем Уиттеридж стал включать световой стимул и регистрировать металлическим электродом электрическую активность с поверхности коры. Он действительно получил в своем опыте ответы, однако они возникали только на внутренней границе поля 17, т.е. в зоне, получающей входные сигналы от длинной, узкой вертикальной полоски в середине поля зрения: при стимуляции маленькими пятнышками света ответы появлялись только тогда, когда свет вспыхивал на вертикальной средней линии или поблизости от нее. Если кору противоположного полушария охлаждали, тем самым временно подавляя ее функцию, ответы прекращались; к этому же приводило и охлаждение мозолистого тела. Тогда стало ясно, что мозолистое тело не может связывать всё поле 17 левого полушария со всем полем 17 правого полушария, а связывает только небольшие участки этих полей, где находятся проекции вертикальной линии в середине поля зрения.
Подобный результат можно было предвидеть исходя из ряда анатомических данных. Только один участок поля 17, расположенный очень близко к границе с полем 18, посылает аксоны через мозолистое тело в другое полушарие, и большая часть их, по-видимому, оканчивается в поле 18 около границы с полем 17. Если мы предположим, что входы в кору от НКТ точно соответствуют контралатеральным частям поля зрения (а именно левое полуполе отображается в коре правого полушария, а правое — в коре левого), то наличие связей между полушариями через мозолистое тело должно приводить в итоге к тому, что каждое полушарие будет получать сигналы от области несколько большей, чем половина поля зрения. Иными словами, за счет связей через мозолистое тело произойдет перекрывание полуполей, проецируемых в два полушария. Именно это мы и обнаружили. С помощью двух электродов, введенных в область коры у границы полей 17 и 18 в каждом из полушарий, нам нередко удавалось регистрировать активность клеток, рецептивные поля которых взаимно перекрывались на несколько угловых градусов.
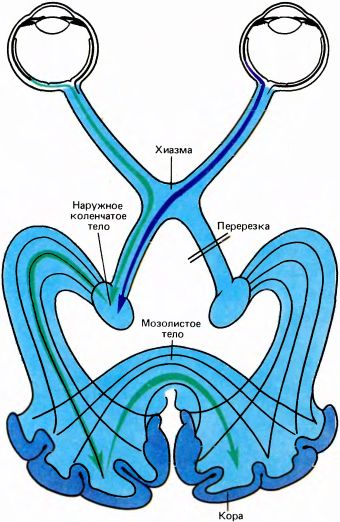
Рис. 100. Уиттеридж в своем эксперименте произвел перерезку правого зрительного тракта. Теперь для того, чтобы информация поступала в зрительную кору правого полушария, она должна сначала попасть в зрительную кору левого полушария, а затем пройти в другое полушарие через мозолистое тело. Охлаждение какого-либо участка этого пути блокирует передачу нервных импульсов.
Т. Визел и я вскоре произвели микроэлектродные отведения непосредственно от той зоны мозолистого тела (в самой задней его части), где имеются волокна, связанные со зрительной системой. Мы нашли, что почти все волокна, которые мы могли активировать зрительными стимулами, отвечали в точности так же, как и обычные нейроны поля 17, т.е. проявляли свойства как простых, так и сложных клеток, избирательно чувствительных к ориентации стимула и обычно отвечавших на стимуляцию обоих глаз. Во всех этих случаях рецептивные поля располагались очень близко к средней вертикали ниже или выше (либо на уровне) точки фиксации, как показано на рис. 101.
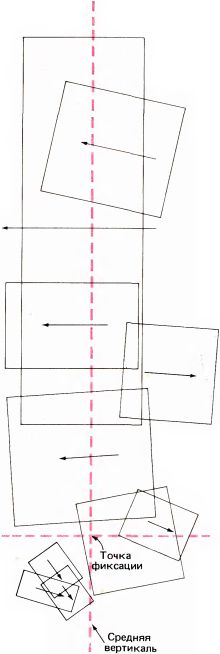
Рис. 101. Рецептивные поля волокон мозолистого тела лежат очень близко к средней вертикали поля зрения. Все эти поля были выявлены путем регистрации активности десяти волокон у одной кошки.
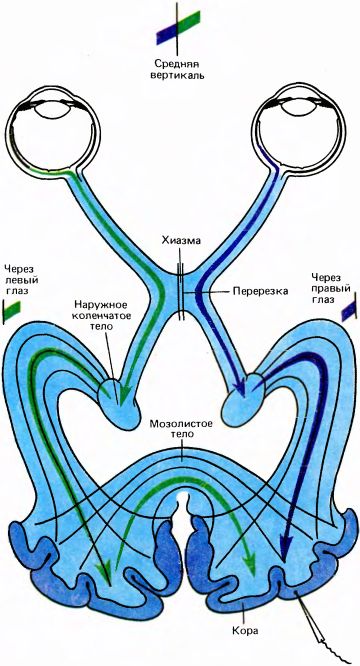
Рис. 102. Берлукки и Риццолатти в своем эксперименте изящно продемонстрировали не только функцию зрительной части волокон мозолистого тела, но и высокую специфичность межполушарных связей между клетками, сходными по ориентации и расположению рецептивных полей. Берлукки и Риццолатти перерезали у кошки хиазму по средней линии, так что левый глаз стал передавать информацию только в левое полушарие, причем эта информация касалась только правой половины поля зрения. Точно так же правый глаз стал снабжать информацией только правое полушарие, и эта информация относилась только к левой половине поля зрения. После перерезки регистрировались ответы тех клеток, рецептивные поля которых в нормальных условиях пересекали бы среднюю вертикаль. Оказалось, что рецептивные поля таких клеток расщепляются по этой вертикали; правая часть связана теперь с левым глазом, а левая — с правым глазом.
Пожалуй, наиболее изящной нейрофизиологической демонстрацией роли мозолистого тела стала работа Дж. Берлукки и Дж. Риццолатти из г. Пизы, выполненная в 1968 году. Перерезав зрительную хиазму по средней линии, они регистрировали ответы в поле 17 вблизи границы с полем 18, отыскивая те клетки, которые могли активироваться бинокулярно. Ясно, что любая бинокулярная клетка этой области в правом полушарии должна получать входные сигналы как прямо от правого глаза (через НКТ), так и от левого глаза и левого полушария через мозолистое тело. Как выяснилось, рецептивное поле каждой бинокулярной клетки захватывало среднюю вертикаль сетчатки, причем та его часть, которая относится к левой половине поля зрения, доставляла информацию от правого глаза, а та, которая заходит в правую половину, — от левого глаза. Другие свойства клеток, исследованные в этом эксперименте, включая ориентационную избирательность, оказались идентичными (рис. 102).
Полученные результаты ясно показали, что мозолистое тело связывает клетки друг с другом таким образом, чтобы их рецептивные поля могли заходить и вправо, и влево от средней вертикали. Таким образом, оно как бы склеивает две половины изображения окружающего мира. Чтобы лучше себе это представить, предположим, что изначально кора нашего мозга образовалась как одно целое, не разделенное на два полушария. В этом случае поле 17 имело бы вид одного непрерывного слоя, на который отображалось бы все зрительное поле. Тогда соседние клетки для реализации таких свойств как, например, чувствительность к движению и ориентационная избирательность, должны были бы иметь, разумеется, сложную систему взаимных связей. Теперь вообразим, что «конструктор» (будь то бог, или, скажем, естественный отбор) решил, что так дальше оставлять нельзя — отныне половина всех клеток должна образовать одно полушарие, а другая половина — другое полушарие. Что тогда нужно сделать со всем множеством межклеточных связей, если две совокупности клеток должны теперь отодвинуться друг от друга? По-видимому, можно просто растянуть эти связи, образовав из них часть мозолистого тела. Для того чтобы устранить задержку при передаче сигналов по такому длинному пути (у человека примерно 12–15 сантиметров), нужно увеличить скорость передачи, снабдив волокна миелиновой оболочкой. Разумеется, на самом деле ничего такого в процессе эволюции не происходило; задолго до того, как возникла кора, мозг уже имел два отдельных полушария.
Эксперимент Берлукки и Риццолатти, на мой взгляд, дал одно из наиболее ярких подтверждений удивительной специфичности нейронных связей. Клетка, показанная на рис. 108 (около кончика электрода) и, вероятно, миллион других подобных клеток, соединенных через мозолистое тело, приобретают свою ориентационную избирательность как за счет локальных связей с соседними клетками, так и за счет связей, идущих через мозолистое тело из другого полушария от клеток с такой же ориентационной чувствительностью и сходным расположением рецептивных полей (сказанное относится и к другим свойствам клеток, таким как дирекциональная специфичность, способность реагировать на концы линий, а также сложность). Каждая из клеток зрительной коры, имеющих связи через мозолистое тело, должна получать входные сигналы от клеток другого полушария с точно такими же свойствами. Мы знаем множество фактов, указывающих на избирательность соединений в нервной системе, но я думаю, что данный пример — наиболее яркий и убедительный.
Рассмотренные выше аксоны клеток зрительной коры составляют лишь небольшую долю всех волокон мозолистого тела. На соматосенсорной коре проводились эксперименты с использованием аксонного транспорта, аналогичные описанным в предыдущих главах опытам с инъекцией радиоактивной аминокислоты в глаз. Их результаты показывают, что мозолистое тело таким же образом связывает те участки коры, которые активируются кожными и суставными рецепторами, расположенными вблизи средней линии тела на туловище и голове, но не связывает корковые проекции конечностей.
Каждая область коры соединяется с несколькими или даже многими другими областями коры того же полушария. Например, первичная зрительная кора связана с полем 18 (зрительной зоной 2), с медиальной височной областью (зоной MT), со зрительной зоной 4 и еще с одной или двумя областями. Многие участки коры имеют также связи с несколькими областями другого полушария, осуществляемые через мозолистое тело, а в некоторых случаях — через переднюю комиссуру. Поэтому мы можем рассматривать эти комиссуральные связи просто как особый вид кортико-кортикальных связей. Легко сообразить, что об этом свидетельствует такой простой пример: если я говорю вам, что моя левая рука ощущает холод или что я увидел что-то слева, то я формулирую слова, используя свои корковые речевые зоны, находящиеся в левом полушарии (сказанное, может быть, и не совсем верно, поскольку я левша); информация, поступающая от левой половины поля зрения или от левой руки, передается в мое правое полушарие; потом соответствующие сигналы должны быть переданы через мозолистое тело в речевую зону коры другого полушария, чтобы я мог сказать что-нибудь о своих ощущениях. В серии работ, начатых в начале 1960-х годов, Р. Сперри (сейчас он работает в Калифорнийском технологическом институте) и его сотрудники показали, что человек с перерезанным мозолистым телом (для лечения эпилепсии) теряет способность рассказывать о тех событиях, информация о которых попадает в правое полушарие. Работа с такими испытуемыми стала ценным источником новых сведений о различных функциях коры, включая мышление и сознание. Первые статьи об этом появились в журнале Brain; они чрезвычайно интересны, и их сможет без труда понять всякий, кто прочел настоящую книгу.
Стереоскопическое зрение
Механизм оценки удаленности, основанный на сравнении двух сетчаточных изображений, настолько надежен, что многие люди (если они не психологи и не специалисты по физиологии зрения) даже не подозревают о его существовании. Для того чтобы убедиться в важности этого механизма, попробуйте в течение нескольких минут вести автомобиль или велосипед, играть в теннис или прокатиться на лыжах, закрыв один глаз. Стереоскопы вышли из моды, и вы можете найти их только в антикварных магазинах. Однако большинство читателей смотрели стереоскопические фильмы (когда зрителю приходится надевать специальные очки). Принцип действия как стереоскопа, так и стереоскопических очков основан на использовании механизма стереопсиса.
Изображения на сетчатках двумерны, а между тем мы видим мир трехмерным. Очевидно, что как для человека, так и для животных важна способность определять расстояние до объектов. Точно так же восприятие трехмерной формы предметов означает оценку относительной глубины. Рассмотрим в качестве простого примера круглый предмет. Если он расположен наклонно по отношению к линии взора, его изображение на сетчатках будет эллиптическим, однако обычно мы без труда воспринимаем такой предмет как круглый. Для этого необходима способность к восприятию глубины.
Человек обладает многими механизмами оценки глубины. Некоторые из них столь очевидны, что вряд ли заслуживают упоминания. Тем не менее я их упомяну. Если приблизительно известна величина объекта, например в случае таких объектов, как человек, дерево или кошка, то можно оценить расстояние до него (правда, есть риск ошибиться, если мы столкнемся с карликом, карликовым деревом или львом). Если один предмет расположен впереди другого и частично его заслоняет, то мы воспринимаем передний объект как расположенный ближе. Если взять проекцию параллельных линий, например железнодорожных рельсов, уходящих вдаль, то в проекции они будут сближаться. Это пример перспективы — весьма эффективного показателя глубины. Выпуклый участок стены кажется более светлым в верхней своей части, если источник света расположен выше (обычно источники света и находятся вверху), а углубление в ее поверхности, если оно освещается сверху, кажется в верхней части более темным. Если же источник света поместить внизу, то выпуклость будет выглядеть как углубление, а углубление — как выпуклость. Важным признаком удаленности служит параллакс движения — кажущееся относительное смещение близких и более далеких предметов, если наблюдатель будет двигать головой влево и вправо или вверх и вниз. Если какой-то твердый предмет поворачивается, пусть даже на небольшой угол, то сразу же выявляется его трехмерная форма. Если мы фокусируем хрусталик нашего глаза на близко расположенном предмете, то более удаленный предмет будет не в фокусе; таким образом, меняя форму хрусталика, т.е. изменяя аккомодацию глаза (см. гл. 2 и 6), мы получаем возможность оценивать удаленность предметов. Если изменять относительное направление осей обоих глаз, сводя их или разводя (осуществляя конвергенцию или дивергенцию), то можно свести вместе два изображения предмета и удерживать их в этом положении. Таким образом, управляя либо хрусталиком, либо положением глаз, можно оценить удаленность объекта. На этих принципах основаны конструкции ряда дальномеров. За исключением конвергенции и дивергенции, все остальные показатели удаленности, перечисленные до сих пор, являются монокулярными. Наиболее важный механизм восприятия глубины — стереопсис — зависит от совместного использования двух глаз. При рассматривании любой трехмерной сцены два глаза формируют несколько различные изображения на сетчатке. Вы легко можете в этом убедиться, если будете смотреть прямо вперед и быстро перемещать голову из стороны в сторону примерно на 10 см или же быстро закрывать поочередно то один, то другой глаз. Если перед вами плоский объект, вы не заметите особой разницы. Однако, если сцена включает предметы на разном расстоянии от вас, вы заметите существенные изменения в картине. В процессе стереопсиса мозг сравнивает изображения одной и той же сцены на двух сетчатках и с большой точностью оценивает относительную глубину.
Предположим, наблюдатель фиксирует взором некоторую точку P. Это утверждение эквивалентно тому, как если мы скажем: глаза направляются таким образом, чтобы изображения точки оказались в центральных ямках обоих глаз (F на рис. 103). Предположим теперь, что Q — это другая точка пространства, которая кажется наблюдателю расположенной на такой же глубине, что и P. Пусть QL и QR — изображения точки Q на сетчатках левого и правого глаза. В этом случае точки QL и QR называют корреспондирующими точками двух сетчаток. Очевидно, что две точки, совпадающие с центральными ямками сетчаток, будут корреспондирующими. Из геометрических соображений ясно также, что точка Q', оцениваемая наблюдателем как расположенная ближе, чем Q, будет давать на сетчатках две проекции — Q'L и Q'R — в некорреспондирующих точках, расположенных дальше друг от друга, чем в том случае, если бы эти точки были корреспондирующими (эта ситуация изображена в правой части рисунка). Точно так же, если рассматривать точку, расположенную дальше от наблюдателя, то окажется, что ее проекции на сетчатках будут расположены ближе друг к другу, чем корреспондирующие точки. То, что сказано выше о корреспондирующих точках, — это частично определения, а частично утверждения, вытекающие из геометрических соображений. При рассмотрении этого вопроса учитывается также психофизиология восприятия, поскольку наблюдатель субъективно оценивает, дальше или ближе точки P расположен объект. Введем еще одно определение. Все точки, которые, подобно точке Q (и, конечно, точке P), воспринимаются как равноудаленные, лежат на гороптере — поверхности, проходящей через точки P и Q, форма которой отличается как от плоскости, так и от сферы и зависит от нашей способности оценивать удаленность, т.е. от нашего мозга. Расстояния от центральной ямки F до проекций точки Q (QL и QR) близки, но не равны. Если бы они всегда были равны, то линия пересечения гороптера с горизонтальной плоскостью представляла бы собой круг.
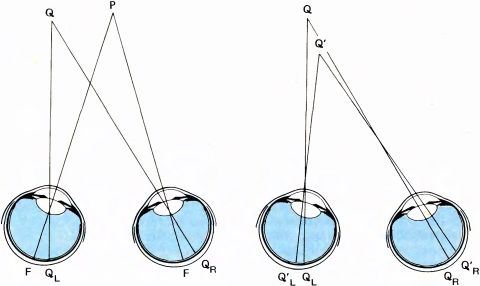
Рис. 103. Слева: если наблюдатель смотрит на точку P, то два ее изображения (проекции) попадают на центральные ямки двух глаз (точки F). Q — точка, которая, по оценке наблюдателя, находится на таком же расстоянии от него, что и P. В этом случае говорят, что две проекции точки Q (QL и QR) попадают в корреспондирующие точки сетчаток. (Поверхность, составленную из всех точек Q, которые кажутся находящимися на одинаковом расстоянии от наблюдателя, таком же, как точка P, называют гороптером, проходящим через точку P). Справа: если точка Q' находится ближе к наблюдателю, чем Q, то ее проекции на сетчатках (Q'L и Q'R) будут отстоять друг от друга по горизонтали дальше, чем если бы они находились в корреспондирующих точках. Если бы точка Q' находилась дальше, то проекции Q'L и Q'R оказались бы сдвинутыми по горизонтали ближе друг к другу.
Предположим теперь, что мы фиксируем взглядом некоторую точку в пространстве и что в этом пространстве расположены два точечных источника света, которые дают проекцию на каждой сетчатке в виде световой точки, причем эти точки — не корреспондирующие: расстояние между ними несколько больше, чем между корреспондирующими точками. Любое такое отклонение от положения корреспондирующих точек мы будем называть диспаратностью. Если это отклонение в горизонтальном направлении не превышает 2° (0,6 мм на сетчатке), а по вертикали не больше нескольких угловых минут, то мы будем зрительно воспринимать одиночную точку в пространстве, расположенную ближе, чем та, которую мы фиксируем. Если же расстояния между проекциями точки будут не больше, а меньше, чем между корреспондирующими точками, то данная точка будет казаться расположенной дальше, чем точка фиксации. Наконец, в том случае, если вертикальное отклонение будет превышать несколько угловых минут или же горизонтальное будет больше 2°, то мы увидим две отдельные точки, которые, возможно, покажутся расположенными дальше или ближе точки фиксации. Эти экспериментальные результаты иллюстрируют основной принцип стереовосприятия, впервые сформулированный в 1838 году сэром Ч. Уитстоном (который также изобрел прибор, известный в электротехнике как «мостик Уитстона»).
Кажется почти невероятным, что до этого открытия ни один человек, по-видимому, не отдавал себе отчета в том, что наличие едва заметных различий в изображениях, проецируемых на сетчатки двух глаз, может приводить к отчетливому впечатлению глубины. Такой стереоэффект может продемонстрировать за несколько минут любой человек, способный произвольно сводить или разводить оси своих глаз, или же тот, у кого есть карандаш, кусок бумаги и несколько небольших зеркал или призм. Непонятно, как прошли мимо этого открытия Евклид, Архимед и Ньютон. В своей статье Уитстон отмечает, что Леонардо да Винчи был очень близок к открытию этого принципа. Леонардо указывал, что шар, расположенный перед какой-либо пространственной сценой, виден каждым глазом по-разному — левым глазом мы немного дальше видим его левую сторону, а правым глазом — правую. Далее Уитстон отмечает, что если бы вместо шара Леонардо выбрал куб, то он, безусловно, заметил бы, что его проекции для разных глаз различны. После этого он мог бы, как и Уитстон, заинтересоваться тем, что будет, если специально спроецировать два подобных изображения на сетчатки двух глаз.
Важным физиологическим фактом является то, что ощущение глубины (т.е. возможность «непосредственно» видеть, дальше или ближе точки фиксации расположен тот или иной объект) возникает в тех случаях, когда два сетчаточных изображения несколько смещены относительно друг друга в горизонтальном направлении — раздвинуты или, наоборот, сближены (если только это смещение не превышает примерно 2°, а вертикальное смещение близко к нулю). Это, разумеется, соответствует геометрическим соотношениям: если по отношению к некоторой точке отсчета расстояния объект расположен ближе или дальше, то его проекции на сетчатках будут раздвинуты или сближены по горизонтали, тогда как существенного вертикального смещения изображений не произойдет.
На этом и основано действие стереоскопа, изобретенного Уитстоном. Стереоскоп в течение примерно полувека был настолько популярен, что имелся чуть ли не в каждом доме. Тот же принцип лежит в основе и стереокино, которое мы сейчас смотрим, используя для этого специальные поляроидные очки. В первоначальной конструкции стереоскопа наблюдатель рассматривал два изображения, помещенные в ящик, с помощью двух зеркал, которые были расположены таким образом, что каждый глаз видел только одно изображение. Для удобства теперь часто используют призмы и фокусирующие линзы. Два изображения идентичны во всем, кроме небольших горизонтальных смещений, которые и создают впечатление глубины. Любой может изготовить фотографию, пригодную для использования в стереоскопе, если выберет какой-либо неподвижный объект (или сцену), сделает снимок, а затем сдвинет фотоаппарат на 5 сантиметров вправо или влево и сделает второй снимок.
Не все обладают способностью воспринимать глубину с помощью стереоскопа. Вы сами можете легко проверить свой стереопсис, если воспользуетесь стереопарами, приведенными на рис. 105 и 106. Если у вас есть стереоскоп, вы можете сделать копии изображенных здесь стереопар и вставить их в стереоскоп. Вы можете также поместить тонкий кусок картона перпендикулярно между двумя изображениями из одной стереопары и попытаться смотреть каждым глазом на свое изображение, установив глаза параллельно, как если бы вы смотрели вдаль. Можно также научиться сводить и разводить глаза с помощью пальца, поместив его между глазами и стереопарой и передвигая вперед или назад, пока изображения не сольются, после чего (это самое трудное) вы сможете рассматривать слитое изображение, стараясь, чтобы оно не разделилось на два. Если у вас это получится, то кажущиеся отношения глубины будут противоположны тем, которые воспринимаются при использовании стереоскопа.

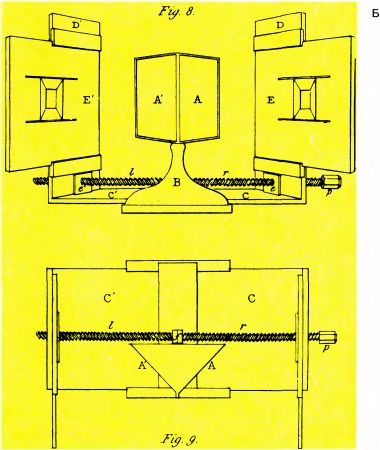
Рис. 104. А. Стереоскоп Уитстона. Б. Схема стереоскопа Уитстона, составленная им самим. Наблюдатель сидит перед двумя зеркалами (А и А'), поставленными под углом 40° к направлению его взора, и смотрит на две совмещенные в поле зрения картинки — Е (правым глазом) и Е' (левым глазом). В созданном позже более простом варианте две картинки помещаются рядом так, что расстояние между их центрами примерно равно расстоянию между глазами. Две призмы отклоняют направление взора так, что при надлежащей конвергенции левый глаз видит левое изображение, а правый глаз — правое изображение. Вы сами можете попробовать обойтись без стереоскопа, представив себе, что смотрите на очень удаленный предмет глазами, оси которых установлены параллельно друг другу. Тогда левый глаз будет смотреть на левое изображение, а правый — на правое.
Даже если вам не удастся повторить опыт с восприятием глубины — из-за того ли, что у вас нет стереоскопа, или потому, что вы не можете произвольно сводить и разводить оси глаз, — вы все-таки сможете понять суть дела, хотя не получите удовольствия от стереоэффекта.
В верхней стереопаре на рис. 105 в двух квадратных рамках имеется по небольшому кружку, один из которых смещен немного влево от центра, а другой — немного вправо. Если рассматривать эту стереопару двумя глазами, используя стереоскоп или иной метод совмещения изображений, то вы увидите кружок не в плоскости листа, а впереди него на расстоянии около 2,5 см. Если так же рассматривать нижнюю стереопару на рис. 105, то кружок будет виден позади плоскости листа. Вы воспринимаете положение кружка таким образом потому, что на сетчатки ваших глаз попадает в точности такая же информация, как если бы кружок действительно находился впереди или позади плоскости рамки.
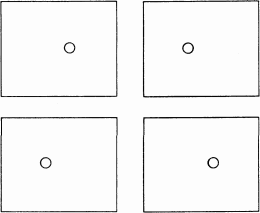
Рис. 105. Если верхнюю стереопару вставить в стереоскоп, то кружок будет выглядеть расположенным впереди плоскости рамки. В нижней стереопаре он будет располагаться позади плоскости рамки. (Вы можете проделать такой опыт без стереоскопа, путем конвергенции или дивергенции глаз; для большинства людей конвергенция легче. Для облегчения задачи можно взять кусок картона и поставить его между двумя изображениями стереопары. Поначалу это упражнение может показаться вам трудным и утомительным; не усердствуйте при первой попытке. При конвергенции глаз на верхней стереопаре кружочек будет виден дальше плоскости, а на нижней — ближе).
В 1960 году Бела Юлеш из фирмы Bell Telephone Laboratories придумал весьма полезную и изящную методику для демонстрации стереоэффекта. Изображение, представленное на рис. 107, на первый взгляд кажется однородной случайной мозаикой из маленьких треугольничков. Так оно и есть, за исключением того, что в центральной части имеется скрытый треугольник большего размера. Если вы будете рассматривать это изображение с помощью двух кусочков цветного целлофана, помещенных перед глазами, — красного перед одним глазом и зеленого перед другим, то вы должны увидеть в центре треугольник, выступающий из плоскости листа вперед, как в предыдущем случае с маленьким кружком на стереопарах. (Быть может, в первый раз вам придется смотреть минуту или около этого, пока не возникнет стереоэффект.) Если поменять куски целлофана местами, произойдет инверсия глубины. Ценность этих стереопар Юлеша заключается в том, что если у вас нарушено стереовосприятие, то вы не увидите треугольника впереди или позади окружающего фона.
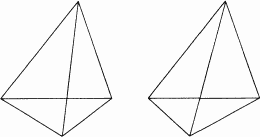
Рис. 106. Еще одна стереопара.
Подводя итоги, можно сказать, что наша способность ощущать стереоэффект зависит от пяти условий:
1. Имеется много косвенных признаков глубины — частичное заслонение одних предметов другими, параллакс движения, вращение предмета, относительные размеры, отбрасывание теней, перспектива. Однако наиболее мощным механизмом является стереопсис.
2. Если мы фиксируем взглядом какую-то точку в пространстве, то проекции этой точки попадают в центральные ямки обеих сетчаток. Любая точка, которая оценивается как расположенная на том же расстоянии от глаз, что и точка фиксации, образует две проекции в корреспондирующих точках сетчаток.
3. Стереоэффект определяется простым геометрическим фактом — если некоторый объект находится ближе точки фиксации, то две его проекции на сетчатках оказываются дальше друг от друга, чем корреспондирующие точки.
4. Главный вывод, основанный на результатах экспериментов с испытуемыми, заключается в следующем: объект, проекции которого на сетчатках правого и левого глаза попадают на корреспондирующие точки, воспринимается как расположенный на том же расстоянии от глаз, что и точка фиксации; если проекции этого объекта раздвинуты по сравнению с корреспондирующими точками, объект кажется расположенным ближе точки фиксации; если же они, наоборот, сближены, объект кажется расположенным дальше точки фиксации.
5. При горизонтальном смещении проекций больше чем на 2° или вертикальном смещении больше нескольких угловых минут возникает двоение.
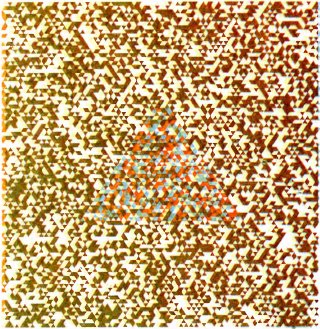
Рис. 107. Для того чтобы получить это изображение, называемое анаглифом, Бела Юлес сначала построил две системы случайно расположенных маленьких треугольников; они различались только тем, что 1) в одной системе были красные треугольники на белом фоне, а в другой — зеленые на белом фоне; 2) в пределах большой треугольной зоны (вблизи центра рисунка) все зеленые треугольники несколько смещены влево по сравнению с красными. После этого две системы совмещаются, но с небольшим сдвигом, так что сами треугольники не накладываются друг на друга. Если на полученное изображение смотреть через зеленый фильтр из целлофана, то будут видны только красные элементы, а если через красный фильтр — только зеленые. Если же перед одним глазом поместить зеленый фильтр, а перед другим — красный, то вы увидите большой треугольник, выступающий примерно на 1 см перед страницей. В случае перемены фильтров местами треугольник будет виден за плоскостью страницы.
Физиология стереоскопического зрения
Если мы хотим знать, каковы мозговые механизмы стереопсиса, то проще всего начать с вопроса: существуют ли нейроны, реакции которых специфически определяются относительным горизонтальным смещением изображений на сетчатках двух глаз? Посмотрим сначала, как отвечают клетки нижних уровней зрительной системы при одновременной стимуляции обоих глаз. Мы должны начать с нейронов поля 17 или более высокого уровня, поскольку ганглиозные клетки сетчатки явно монокулярные, а клетки наружного коленчатого тела, в котором входы от правого и левого глаз распределены по разным слоям, тоже можно считать монокулярными — они отвечают на стимуляцию либо одного глаза, либо другого, но не обоих одновременно. В поле 17 примерно половину нейронов составляют бинокулярные клетки, отвечающие на стимуляцию обоих глаз. При тщательном тестировании выясняется, что ответы этих клеток, по-видимому, мало зависят от относительного положения проекций стимулов на сетчатках двух глаз. Рассмотрим типичную сложную клетку, которая отвечает непрерывным разрядом на движение стимульной полосы через ее рецептивное поле в том или другом глазу. При одновременной стимуляции обоих глаз частота разрядов этой клетки выше, чем при стимуляции одного глаза, но обычно для ответа такой клетки несущественно, попадают ли в какой-то момент проекции стимула точно в одни и те же участки двух рецептивных полей. Наилучший ответ регистрируется тогда, когда эти проекции входят и выходят из соответствующих рецептивных полей двух глаз примерно в одно время; однако не столь важно, которая из проекций немного опережает другую. На рис. 108 показана характерная кривая зависимости ответа (например, общего числа импульсов в ответе за одно прохождение стимула через рецептивное поле) от разницы в положении стимула на обеих сетчатках. Эта кривая очень близка к горизонтальной прямой, из чего ясно, что относительное положение стимулов на двух сетчатках не очень существенно. Клетка такого типа будет хорошо реагировать на линию надлежащей ориентации независимо от ее удаленности — расстояние до линии может быть больше, равно или меньше расстояния до точки, фиксируемой взором.

Рис. 108. При одновременной стимуляции обоих глаз вертикальной светлой линией, движущейся влево, обычная бинокулярная клетка в поле 17 будет давать одинаковые ответы при трех разных относительных положениях этой линии на обеих сетчатках. Нулевая диспаратность означает, что различия в положении нет, — как если бы обезьяна смотрела на экран, на котором предъявляются стимулы. Отсутствие диспаратности не приводит к заметному усилению ответа клетки.
По сравнению с этой клеткой нейроны, ответы которых представлены на рис. 109 и 110, весьма чувствительны к относительному положению двух стимулов на двух сетчатках, т.е. чувствительны к глубине. Первый нейрон (рис. 109) лучше всего отвечает в том случае, если стимулы попадают точно на корреспондирующие участки двух сетчаток. Величина несовмещения стимулов по горизонтали (т.е. диспаратность), при которой клетка уже перестает реагировать, составляет некоторую долю ширины ее рецептивного поля. Поэтому клетка отвечает тогда и только тогда, когда объект находится примерно на таком же расстоянии от глаз, как и точка фиксации. Второй нейрон (рис. 110) отвечает только тогда, когда объект расположен дальше точки фиксации. Имеются также клетки, отвечающие только тогда, когда стимул расположен ближе этой точки. При изменении степени диспаратности нейроны двух последних типов, называемые дальними клетками и ближними клетками, очень резко изменяют интенсивность своих ответов в точке нулевой диспаратности или поблизости он нее. Нейроны всех трех типов (клетки, настроенные на диспаратность) были обнаружены в поле 17 обезьяны. Пока не совсем ясно, насколько часто они там встречаются, расположены ли они в определенных слоях коры и находятся ли в определенных пространственных отношениях к колонкам глазодоминантности. Эти клетки весьма чувствительны к расстоянию объекта от глаз, которое кодируется в виде относительного положения соответствующих стимулов на двух сетчатках. Еще одна особенность этих клеток — то, что они не отвечают на стимуляцию только одного глаза или же отвечают, но очень слабо. Все эти клетки обладают общим свойством ориентационной избирательности; насколько нам известно, они сходны с обычными сложными клетками верхних слоев коры, но обладают еще дополнительным свойством — чувствительностью к глубине. Кроме того, эти клетки хорошо реагируют на движущиеся стимулы, а иногда и на концы линий.

Рис. 109. В ответе этой избирательно отвечающей клетки наблюдается значительная разница в зависимости от того, находится ли стимул точно на том же расстоянии, что и точка фиксации, или же он расположен ближе или дальше. Данная клетка отвечает только тогда, когда расстояние до стимула такое же, как и до точки фиксации. В этом эксперименте ось одного глаза отклоняли в горизонтальном направлении с помощью призмы; однако к такому же результату привело бы и смещение экрана по направлению к животному или от него.
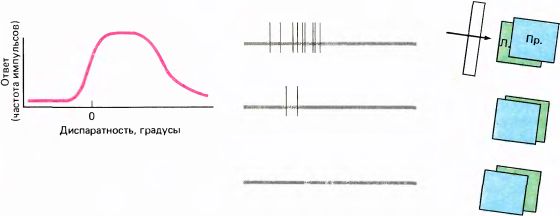
Рис. 110. У клетки представленного здесь типа («дальней») объекты, расположенные ближе плоскости экрана, вызывают очень слабый ответ или вовсе его не вызывают. При нулевой диспаратности (т.е. при расстоянии до стимула, равном расстоянию до экрана) небольшое смещение экрана сильно влияет на реакцию: она резко возрастает и остается постоянной для стимулов, расположенных дальше точки фиксации. Если отвести стимул достаточно далеко, то два рецептивных поля клетки уже не перекрываются, т.е., по существу, два глаза стимулируются независимо. Ответ в этом случае прекращается.
Дж. Поджо из медицинской школы Джонса Гопкинса регистрировал ответы таких клеток в поле 17 бодрствующей обезьяны с вживленными электродами, которая была предварительно обучена фиксировать взглядом определенный объект. У наркотизированных обезьян такие клетки тоже выявлялись в коре, но редко встречались в поле 17 и очень часто — в поле 18. Я был бы крайне удивлен, если бы оказалось, что животные и человек могут стереоскопически оценивать расстояния до объектов с помощью только трех описанных выше типов клеток — настроенных на нулевую диспаратность, «ближних» и «дальних». Я бы скорее ожидал найти полный набор клеток для всех возможных глубин. У бодрствующих обезьян Поджо встречал также узконастроенные клетки, которые лучше всего реагировали не на нулевую диспаратность, а на небольшие отклонения от нее; по-видимому, в коре могут быть специфические нейроны для всех ступеней диспаратности. Хотя мы до сих пор не знаем, как именно мозг «реконструирует» сцену, включающую множество разноудаленных объектов (что бы мы ни понимали под словом «реконструкция»), клетки вроде описанных выше, вероятно, участвуют в первых этапах этого процесса.
Некоторые проблемы, связанные со стереоскопическим зрением
За время изучения стереопсиса психофизики столкнулись с целым рядом проблем. Оказалось, что обработка некоторых бинокулярных стимулов происходит в зрительной системе совершенно непонятными способами. Я мог бы привести много примеров такого рода, но ограничусь лишь двумя.
На примере стереопар, приведенных на рис. 105, мы видели, что смещение двух идентичных изображений (в данном случае кружков) по направлению друг к другу приводит к ощущению большей близости, а по направлению друг от друга — к ощущению большей удаленности. Предположим теперь, что мы делаем одновременно обе эти операции, для чего помещаем в каждую рамку по два кружка, расположенных друг около друга (рис. 111). Очевидно, что рассматривание такой стереопары могло бы привести к восприятию двух кружков — одного ближе, а другого дальше плоскости фиксации. Однако можно предположить и другой вариант: мы увидим просто два кружка, лежащие рядом в плоскости фиксации. Дело в том, что этим двум пространственным ситуациям соответствуют одинаковые изображения на сетчатках. В действительности данная пара стимулов может быть воспринята только как два кружка в плоскости фиксации, в чем легко убедиться, если любым способом достичь слияния квадратных рамок на рис. 111. Точно так же можно представить себе ситуацию, когда мы рассматриваем две цепочки из знаков ?, скажем, по шесть знаков в цепочке. Если рассматривать их в стереоскоп, то в принципе можно воспринять любую из ряда возможных конфигураций в зависимости от того, какой знак ? из левой цепочки сольется с определенным знаком ? в правой цепочке. На самом же деле, если мы будем рассматривать такую стереопару в стереоскоп (или иным способом, создающим стереоэффект), то всегда увидим шесть знаков ? в плоскости фиксации. Мы до сих пор не знаем, как мозг разрешает эту неоднозначность и выбирает простейшую из всех возможных комбинаций. Из-за такого рода неоднозначностей трудно даже вообразить, как нам удается воспринимать объемной сцену, включающую множество ветвей разной величины, находящихся на разных расстояниях от нас. Правда, физиологические данные подсказывают, что задача, возможно, не столь трудна, так как разные ветви скорее всего будут иметь разную ориентацию, а мы уже знаем, что клетки, участвующие в стереопсисе, всегда бывают ориентационно-избирательными.

Рис. 111. Стереопары, приведенные на рис. 105, вызывали ощущение, что кружок расположен ближе или дальше плоскости рамки. Здесь эти две стереопары объединены, и поэтому, казалось бы, мы должны видеть один кружок ближе, а другой дальше. Однако на самом деле этого не получается; оба кружка видны на том же расстоянии, что и рамка.
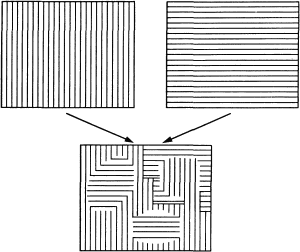
Рис. 112. Эту стереопару невозможно слить в единое восприятие, как другие стереопары (например, показанные на рис. 105 и 106). Вместо этого получается эффект «борьбы полей зрения» — вы видите мозаичную картину, состоящую из фрагментов обоих изображений и беспрестанно меняющуюся.
Второй пример непредсказуемости бинокулярных эффектов, имеющий отношение к стереопсису, — это так называемая борьба полей зрения, о которой мы упоминаем также в разделе о косоглазии (гл. 9). Если на сетчатках правого и левого глаза создаются очень сильно различающиеся изображения, то часто одно из них перестает восприниматься. Если вы будете смотреть левым глазом на решетку из вертикальных линий, а правым глазом — на решетку из горизонтальных линий (рис. 112; можно пользоваться стереоскопом или конвергенцией глаз), то, казалось бы, следует ожидать, что вы увидите сетку из пересекающихся линий. Однако в действительности почти невозможно увидеть оба набора линий одновременно. Виден или тот или другой, причем каждый из них — лишь в течение нескольких секунд, после чего он исчезает и появляется другой. Иногда можно также увидеть как бы мозаику из двух этих изображений, в которой отдельные однородные участки будут перемещаться, сливаться или разделяться, а ориентация линий в них будет меняться (см. рис. 112, внизу). По какой-то причине нервная система не может воспринимать столь разные стимулы одновременно в одном и том же участке поля зрения, и она подавляет переработку одного из них. Слово «подавлять» мы используем здесь просто как иное описание того же феномена: на самом деле мы не знаем, как осуществляется такое подавление и на каком уровне центральной нервной системы оно происходит. Мне думается, мозаичный характер воспринимаемого образа при борьбе полей зрения позволяет предположить, что «принятие решений» в этом процессе происходит на достаточно ранних этапах переработки зрительной информации, возможно, в поле 17 или 18. (Я рад, что мне не нужно защищать это предположение.)
Феномен борьбы полей зрения означает, что в тех случаях, когда зрительная система не может объединить изображения на двух сетчатках (в плоскую картину, если изображения одинаковы, или в трехмерную сцену, если имеется лишь небольшая горизонтальная диспаратность), она просто отвергает один из образов — либо полностью, когда, например, мы смотрим в микроскоп, держа второй глаз открытым, либо частично или на время, как в примере, описанном выше. В ситуации с микроскопом существенную роль играет внимание, но нейронные механизмы, лежащие в основе такого переключения внимания, тоже неизвестны.
Еще один пример борьбы полей зрения вы можете наблюдать, если просто будете рассматривать какую-нибудь многоцветную сцену или картину через очки с красным и зеленым светофильтрами. Впечатления разных наблюдателей в этом случае могут быть весьма различными, однако большинство людей (в том числе и я) отмечает переходы от общего красноватого тона к зеленоватому и обратно, но без желтого цвета, который получается при обычном смешении красного света с зеленым (см. гл. 8, рис. 121).
Стереослепота
Если человек слеп на один глаз, то очевидно, что он не будет обладать стереоскопическим зрением. Однако его нет и у некоторой части людей, зрение которых в остальных отношениях нормально. Удивительно то, что доля таких людей не слишком мала. Так, если показать стереопары вроде тех, что приводились на рис. 105 и 106, сотне студентов-испытуемых (применяя поляроиды и поляризованный свет), то обычно оказывается, что четыре или пять из них не могут достичь стереоэффекта. Нередко это удивляет их самих, так как в повседневных условиях они не испытывают никаких неудобств. Последнее может показаться странным любому, кто ради эксперимента пытался управлять автомобилем, закрыв один глаз. По-видимому, отсутствие стереопсиса достаточно хорошо компенсируется использованием других признаков глубины, таких как параллакс движения, перспектива, частичное закрытие одних предметов другими и т.п. В главе 9 мы рассмотрим случаи врожденного косоглазия, когда глаза длительное время работают несогласованно. Это может приводить к нарушению связей в коре, обеспечивающих бинокулярное взаимодействие, и в результате — к утрате стереопсиса. Косоглазие встречается не так уж редко, и даже слабая его степень, которая может остаться незамеченной, в некоторых случаях, вероятно, бывает причиной стереослепоты. В других же случаях нарушение стереопсиса, подобно цветовой слепоте, может быть наследственным.
Поскольку в этой главе шла речь и о мозолистом теле, и о стереоскопическом зрении, я воспользуюсь случаем сказать кое-что о связи этих двух вещей. Попробуйте задать себе вопрос: каких нарушений стереопсиса можно ожидать у человека с перерезанным мозолистым телом? Ответ на этот вопрос ясен из схемы, приведенной на рис. 113.
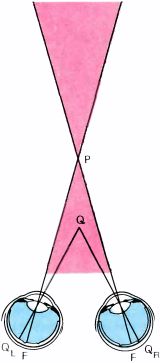
Рис. 113. Перерезка мозолистого тела приводит к потере стереопсиса в закрашенной части видимого пространства.
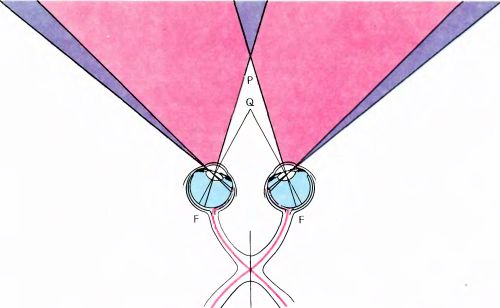
Рис. 114. Результаты продольной перерезки хиазмы по средней линии. Испытуемый совсем не будет видеть две более темные зоны по краям рисунка, слева и справа. Между этими зонами, там, где окраска светлее, не будет стереопсиса, за исключением небольшой зоны треугольной формы за точкой P (здесь вообще ничего не видно) и зоны впереди точки P (здесь стереопсис сохранится).
Если человек фиксирует взглядом точку P, то проекции точки Q, расположенной ближе к глазам в пределах острого угла FPF, — QL и QR — окажутся в левом и в правом глазу по разные стороны от центральной ямки. Соответственно проекция QL передает информацию в левое полушарие, а проекция QR — в правое полушарие. Для того чтобы увидеть, что точка Q ближе, чем P (т.е. получить стереоэффект), нужно объединить информацию левого и правого полушарий. Но единственный способ сделать это — передать информацию по мозолистому телу. Если же путь через мозолистое тело разрушен, человек окажется стереослепым в закрашенной на рисунке области. В 1970 году Д. Митчелл и К. Блейкмор из Калифорнийского университета в Беркли исследовали стереоскопическое зрение у одного человека с перерезанным мозолистым телом и получили в точности предсказанный выше результат.
Второй вопрос, тесно связанный с первым, состоит в том, какое нарушение стереопсиса произойдет, если перерезать по средней линии зрительную хиазму (что проделал Р. Майерс на кошках). Результат здесь будет в определенном смысле противоположным. Из рис. 114 должно быть ясно, что в этом случае каждый глаз станет слепым в отношении стимулов, падающих на носовую область сетчатки, т.е. исходящих из височной части поля зрения. Поэтому стереопсиса не будет в области пространства, окрашенной светлее, где он в норме имеется. Боковые зоны за пределами этой области вообще доступны только для одного глаза, так что стереопсис здесь отсутствует и в нормальных условиях, а после перерезки хиазмы они будут зонами слепоты (на рисунке это показано более темным цветом). В области позади точки фиксации, где перекрываются височные части полей зрения, ставшие теперь невидимыми, тоже наступит слепота. Однако в зоне ближе точки фиксации сохранившиеся полуполя обоих глаз перекрываются, так что здесь должен сохраниться стереопсис, если только не повреждено мозолистое тело. К. Блейкмор нашел все-таки больного с полной перерезкой хиазмы по средней линии (этот больной, будучи ребенком, получил перелом черепа при езде на велосипеде, что, по-видимому, привело к продольному разрыву хиазмы). При проверке у него была обнаружена именно та комбинация дефектов зрения, которую мы только что гипотетически описали.
Глаз, мозг, зрение
Дэвид Хантер Хьюбел; David Hunter Hubel;
Дек 24 2017