
Статьи. Предметно
Цветовое зрение
Цветовое зрение
Сотни дополнительных долларов, которые покупатели соглашаются платить за цветной телевизор, предпочитая его черно-белому, означают, что цветовые ощущения для нас достаточно важны. Сложный аппарат глаза и мозга может воспринимать различия в спектральном составе света, отражаемого от видимых предметов, и легко представить себе, какие преимущества давала эта способность нашим предкам. Одним из преимуществ, несомненно, было то, что она затрудняла маскировку другим животным: потенциальной добыче намного труднее слиться с окружающим фоном, если хищник может различать не только интенсивность света, но и цвет. Столь же важным цвет может быть при поиске растительной пищи: обезьяна легко найдет ярко-красную ягоду, выделяющуюся среди зеленой листвы, и это даст животному несомненное преимущество, как, впрочем, и растению, поскольку семена проходят невредимыми через пищеварительный тракт обезьяны и рассеиваются на обширной площади. Для некоторых животных цвет важен при размножении; примерами служат ярко-красная окраска области промежности у макаков и изумительное оперение у самцов многих птиц.
У людей давление отбора, направленное на сохранение или улучшение цветового зрения, видимо, ослабевает, судя по тому, что 7 или 8 процентов мужчин частично или полностью лишены цветового зрения, но отлично без него обходятся, причем этот дефект часто долгие годы остается незамеченным и выявляется лишь после того, как за рулем они проедут на красный свет. Даже те из нас, кто обладает нормальным цветовым зрением, могут испытывать подлинное наслаждение от черно-белых фильмов, которые в художественном отношении иногда могут быть шедеврами киноискусства. Как мы увидим позже, при слабом освещении все мы в цветовом отношении слепы.
Ощущение цвета у позвоночных встречается спорадически; вероятно, в ходе эволюции оно неоднократно редуцировалось или даже исчезало, чтобы потом появиться снова. К млекопитающим, у которых цветовое зрение слабо развито или отсутствует, относятся мыши, крысы, кролики, кошки, собаки и ночная обезьяна дурукули. У сусликов и приматов, включая людей, человекообразных и большинство других обезьян, цветовое зрение хорошо развито. Из ночных животных, зрение которых приспособлено к слабому свету, лишь немногие хорошо различают цвета; это позволяет думать, что по каким-то причинам различение цветов и способность видеть при слабом свете несовместимы друг с другом. Среди других позвоночных цветовое зрение хорошо развито у многих рыб и птиц, но, вероятно, отсутствует или слабо выражено у рептилий и амфибий. Цветовым зрением обладают многие насекомые, в том числе мухи и пчелы. В отношении подавляющего большинства животных у нас нет точных сведений о способности различать цвета — вероятно, потому, что проводить поведенческие или физиологические тесты на цветовое зрение не так легко.

Рис. 115. Цвет используется в живой природе для разных целей, и некоторые из них пока не известны. Синие пятнышки на боках этой рыбы (Hypsypops) становятся все менее яркими, по мере того как рыба растет, и исчезают, когда она достигает зрелости. Какое значение имеют эти пятнышки для других особей того же вида, неизвестно.
Вопросом о цветовом зрении — несоразмерно его биологическому значению для человека — занимался ряд блестящих умов, включая Ньютона, Гёте (сильной стороной которого не были, однако, естественные науки) и Гельмгольца. Тем не менее до сих пор даже художники, физики и биологи часто плохо представляют себе, что такое цвет. Проблема возникает еще в детстве, когда нам впервые дают коробку с красками, а затем говорят, что желтый, синий и красный — это «основные» цвета и что желтый с синим дают зеленый. Многие из нас впоследствии поражаются кажущемуся противоречию с этим фактом, когда с помощью пары проекторов мы отбрасываем на экран два перекрывающихся пятна, желтое и синее, и видим в области их наложения красивый белый цвет. Результат смешения красок — это предмет физики; смешение же световых лучей — в основном вопрос биологии.
Рассуждая о цвете, полезно мысленно разделять эти два аспекта — физический и биологический. Физика, которую нам при этом следует знать, ограничивается лишь некоторыми фактами о световых волнах. Биология же включает психофизику и физиологию. Психофизику интересуют наши чувства как детекторы внешней информации, а физиологию — лежащие в их основе внутренние механизмы, в частности работа нашей зрительной системы. Мы многое знаем о физике и психофизике цвета, но физиология находится все еще на сравнительно примитивном уровне, главным образом из-за того, что необходимые методы стали доступны лишь в последние десятилетия.
Природа света
Свет состоит из частиц, называемых фотонами, каждую из которых можно рассматривать как пакет электромагнитных волн. Будет ли луч электромагнитной энергии именно светом, а не рентгеновскими лучами или радиоволнами, определяется длиной волны — расстоянием от одного гребня волны до следующего: в случае света это расстояние составляет приблизительно 0,0000001 (10–7) метра, или 0,0005 миллиметра, или 0,5 микрометра, или 500 нанометров (нм).
Свет — это по определению то, что мы можем видеть. Наши глаза могут воспринимать электромагнитные волны длиной от 400 до 700 нм. Обычно попадающий в наши глаза свет состоит из сравнительно однородной смеси лучей с различными длинами волн; такую смесь называют белым светом (хотя это весьма нестрогое понятие). Для оценки волнового состава световых лучей измеряют световую энергию, заключенную в каждом из последовательных небольших интервалов, например от 400 до 410 нм, от 410 до 420 нм и т.д., после чего рисуют график распределения энергии по длинам волн. Для света, приходящего от Солнца, этот график похож на левую кривую на рис. 116. Это кривая без резких подъемов и спадов с пологим максимумом в области 600 нм. Такая кривая типична для излучения раскаленного объекта. Положение максимума зависит от температуры источника: для Солнца это будет область около 600 нм, а для звезды более горячей, чем наше Солнце, максимум сдвинется к более коротким волнам — к голубому концу спектра, т.е. на нашем графике — влево. (Представление художников о том, что красные, оранжевые и желтые цвета — теплые, а синие и зеленые — холодные, связано только с нашими эмоциями и ассоциациями и не имеет никакого отношения к спектральному составу света от раскаленного тела, зависящему от его температуры, — к тому, что физики называют цветовой температурой.)
Если мы будем каким-то способом фильтровать белый свет, удаляя все, кроме узкой спектральной полосы, то получим свет, который называют монохроматическим (см. график на рис. 116 справа).
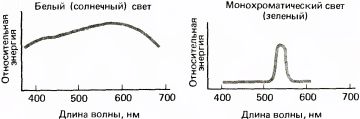
Рис. 116. Слева: энергия света (например, солнечного) распределена в широком диапазоне длин волн — примерно от 400 до 700 нанометров. Слабо выраженный пик определяется температурой источника: чем горячее источник, тем больше смещение пика к синему (коротковолновому) концу. Справа: монохроматический свет — это свет, энергия которого сосредоточена в основном в области какой-то одной длины волны. Его можно создать при помощи разнообразных фильтров, лазера или спектроскопа с призмой или дифракционной решеткой.
Пигменты
Когда свет падает на некоторый объект, может происходить одно из трех событий: свет может поглощаться, а энергия его превращаться в тепло, как это бывает, когда что-то нагревается на солнце; он может проходить сквозь объект, если, например, на пути солнечных лучей окажется вода или стекло; либо он может отражаться, как в случае зеркала или любого светлого предмета, например куска мела. Часто происходят два или все три события; например, часть света может поглотиться, а часть — отразиться. Для многих объектов относительное количество поглощенного и отраженного света зависит от длины волны. Зеленый лист растения поглощает длинно- и коротковолновый свет и отражает свет промежуточной области спектра, так что при освещении листа солнечными лучами отраженный свет будет иметь выраженный широкий максимум на средних длинах волн (в области зеленого цвета). Красный объект будет иметь свой максимум, тоже широкий, в области длинных волн, как показано на рис. 117.
Вещество, которое поглощает часть падающего на него света и отражает остальную часть, называют пигментом. Если какие-то спектральные компоненты в диапазоне видимого света поглощаются лучше, чем другие, пигмент представляется нам окрашенным. Сразу же добавим: какой именно цвет мы видим, зависит не только от длины волн, но также от распределения энергии между разными участками спектра и от свойств нашей зрительной системы. Здесь замешаны как физика, так и биология.
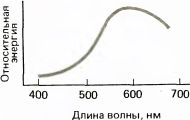
Рис. 117. Большинство окрашенных объектов отражает свет, энергия которого обычно в некоторых участках видимого спектра больше, чем в других участках. Распределение его энергии, однако, намного шире, чем у монохроматического света. На этом графике показан спектральный состав света, который отразится от красного объекта при его освещении широкополосным (белым) источником.
Зрительные рецепторы
Каждая палочка или колбочка в нашей сетчатке содержит пигмент, поглощающий в каком-то участке спектра лучше, чем в других участках. Поэтому, если бы мы смогли собрать достаточное количество такого пигмента и посмотреть на него, он выглядел бы окрашенным. Зрительный пигмент обладает особым свойством: при поглощении им светового фотона он изменяет свою молекулярную форму и при этом высвобождает энергию, запуская таким образом цепь химических реакций, описанную в главе 3, которые в конце концов приводят к появлению электрического сигнала и к выделению химического медиатора в синапсе. Пигментная молекула в своей новой форме, как правило, обладает совсем иными светопоглощающими свойствами, и если, как это обычно бывает, она поглощает свет хуже, чем в исходной форме, мы говорим, что она «выцветает» под действием света. Затем сложный химический механизм глаза восстанавливает первоначальную конфигурацию пигмента; в противном случае его запас быстро истощился бы.
Сетчатка содержит своего рода мозаику из рецепторов четырех типов — палочек и трех типов колбочек (рис. 118). Каждый тип рецепторов содержит свой особый пигмент. Разные пигменты отличаются друг от друга в химическом отношении, а в связи с этим и по способности поглощать свет с различной длиной волн. Палочки ответственны за нашу способность видеть при слабом свете, т.е. за сравнительно грубую разновидность зрения, не позволяющую различать цвета. Палочковый пигмент родопсин обладает наибольшей чувствительностью в области около 510 нм, в зеленой части спектра. Палочки отличаются от колбочек во многих отношениях: они меньше и имеют несколько иное строение, по-иному распределены в разных частях сетчатки и имеют свои особенности в системе связей, образуемых с последующими уровнями зрительного пути. И наконец, по содержащимся в них светочувствительным пигментам три типа колбочек отличаются как друг от друга, так и от палочек.
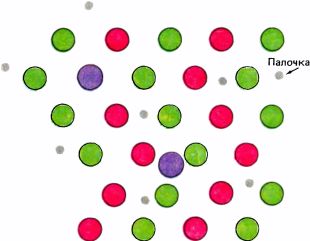
Рис. 118. Рецепторы сетчатки образуют мозаику, состоящую из палочек и трех типов колбочек. Данная схема могла бы отображать участок сетчатки в нескольких градусах от центральной ямки, где колбочек больше, чем палочек.
Пигменты колбочек трех типов имеют пики поглощения в области 430, 530 и 560 нм (рис. 119); поэтому разные колбочки несколько неточно называют соответственно «синими», «зелеными» и «красными». Неточность состоит в том, что 1) эти названия отражают максимумы чувствительности (которые в свою очередь зависят от светопоглощающей способности), а не то, как эти пигменты выглядели бы, если бы на них можно было посмотреть; 2) монохроматический свет с длинами волн 430, 530 и 560 нм будет не синим, зеленым и красным, а фиолетовым, сине-зеленым и желто-зеленым; 3) если бы можно было стимулировать колбочки только одного типа, мы видели бы не синий, зеленый и красный цвета, а, вероятно, фиолетовый, зеленый и желтовато-зеленый. Однако приведенные выше названия колбочек широко распространены, а попытки изменить укоренившуюся терминологию обычно оканчиваются неудачей. Более корректными были бы названия «длинноволновые», «средневолновые» и «коротковолновые», но они затрудняли бы понимание для тех, кто не слишком хорошо знаком со спектром.
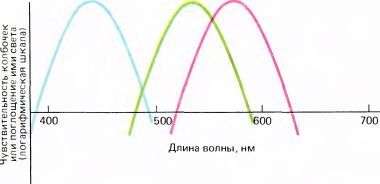
Рис. 119. Спектры поглощения (или кривые спектральной чувствительности) у колбочек трех типов различны. (Ординаты на энергетических кривых и кривых поглощения откладываются в логарифмических единицах, поскольку их значения изменяются в очень широком диапазоне. Поэтому положение оси x произвольно и не соответствует нулевому поглощению).
Имея максимум поглощения в зеленой области, палочковый пигмент родопсин отражает синие и красные лучи и поэтому выглядит пурпурным. Поскольку в наших сетчатках он присутствует в количествах, достаточных для того, чтобы химики смогли его выделить и можно было на него посмотреть, он издавна получил название зрительного пурпура. Само по себе это нелогично, поскольку «зрительный пурпур» называют так по его видимому цвету, тогда как названия для колбочек («красные», «синие» и «зеленые») соответствуют их относительной чувствительности, т.е. способности поглощать свет. Во избежание путаницы об этом следует помнить.
Три типа колбочек имеют широкие зоны чувствительности со значительным перекрыванием, особенно для красных и зеленых колбочек. Свет с длиной волны 600 нм вызовет наибольшую реакцию красных колбочек, пик чувствительности которых расположен при 560 нм; вероятно, он вызовет также некоторую, хотя и более слабую, реакцию колбочек двух других типов. Таким образом, «красная» колбочка реагирует не только на длинноволновый, т.е. красный, свет; она лишь реагирует на него лучше других колбочек. Сказанное относится и к колбочкам других типов.
До сих пор я рассматривал физические аспекты цветового зрения: природу света и пигментов, свойства объектов, отражающих свет к нашим глазам, и особенности палочковых и колбочковых пигментов, преобразующих поглощенный свет в электрические сигналы. Интерпретировать эти исходные сигналы как различные цвета — это уже задача мозга. Чтобы лучше дать почувствовать предмет обсуждения, я решил вначале кратко изложить элементарные факты о цветовом зрении, оставив пока в стороне трехсотлетнюю историю установления этих фактов, а также процессы обработки цветовой информации мозгом.
Общие замечания о цвете
Быть может, полезно начать с того, как оперируют с различными длинами волн две сенсорные системы — слуховая и зрительная. Деятельность одной из них приводит к восприятию высоты тона, а другой — к восприятию цвета, но между этими системами есть глубокое различие. Когда я беру на фортепиано аккорд из пяти нот, вы можете выделить отдельные ноты и пропеть каждую из них по отдельности. Ноты не смешиваются в нашем мозгу, но сохраняют свою индивидуальность, в то время как еще со времен Ньютона известно, что при смешивании двух или нескольких световых лучей разного цвета вы не можете выделить компоненты путем простого рассматривания.
Небольшое размышление убедит вас в том, что цветовое зрение неизбежно должно быть чувством менее совершенным, чем восприятие тонов. Звук, приходящий в любой данный момент в одно ухо и состоящий из колебаний с разной длиной волны, будет воздействовать на тысячи рецепторов внутреннего уха, каждый из которых настроен на высоту, слегка отличающуюся от настройки соседнего рецептора. Если звук состоит из многих волновых составляющих, информацию будет получать множество рецепторов, все выходные сигналы которых передаются в наш мозг. Богатство слуховой информации определяется способностью мозга анализировать такие комбинации звуков.
Совершенно иначе обстоит дело со зрением. Предметом обработки в зрительной системе служит изображение, схватываемое в каждый момент времени набором из миллионов рецепторов. Мы мгновенно воспринимаем сложную сцену. Если при этом мы захотели бы еще обрабатывать длины волн по принципам, используемым во внутреннем ухе, то сетчатка должна была бы иметь не только набор рецепторов, покрывающих всю ее поверхность, но и, скажем, по тысяче рецепторов в каждой отдельной точке, каждый из которых обладал бы максимальной чувствительностью к своей длине волны. Но втиснуть тысячу рецепторов в каждую точку сетчатки физически невозможно; поэтому здесь приходится идти на компромисс. Сетчатка содержит «цветовые» рецепторы трех типов с различной чувствительностью к длине волны в каждой из очень большого числа точек. Таким образом, ценой незначительного ущерба для разрешающей способности бо?льшая часть нашей сетчатки получает некоторую возможность обрабатывать информацию о длинах волн. Мы различаем семь цветов, а не 88 (впрочем, обе цифры следует многократно увеличить с учетом оттенков), но зато каждой из множества тысяч точек видимой сцены будет приписан определенный цвет. Сетчатка не могла бы обладать той способностью к пространственному анализу, которую она имеет, и одновременно обрабатывать информацию о длинах волн столь же изощренно, как слуховая система.
Теперь нужно дать читателю представление о том, что означает для нашего цветового зрения обладание тремя типами колбочек. Во-первых, может возникнуть вопрос: если данная колбочка при каких-то длинах волн работает лучше, чем при других, почему бы зрительной системе просто не измерить выход этой колбочки и не вычислить отсюда, каков здесь цвет? Почему бы тогда не иметь колбочки одного типа вместо трех? Да потому, что при одном типе колбочек, скажем красных, вы не смогли бы отличить свет с наиболее эффективной длиной волны в области 560 нм от более яркого света с менее эффективной длиной волны. Необходимо иметь возможность отличать изменения яркости от изменений длины волны.
Но предположим, что у вас есть два вида колбочек с перекрывающимися кривыми спектральной чувствительности, например красные и зеленые колбочки. Теперь вы можете определять длину волны простым сравнением выходов колбочек. При коротких волнах сильнее будут реагировать зеленые колбочки; по мере увеличения длины волны реакции тех и других колбочек будут все больше приближаться друг к другу, пока не сравняются; примерно при 580 нм красные начнут отвечать лучше зеленых, и эта разница будет постепенно увеличиваться по мере дальнейшего роста длины волны. Если мы вычтем из кривой чувствительности для одних колбочек кривую для других (это логарифмические кривые, поэтому мы фактически берем отношения величин), то мы получим некоторую кривую, не зависящую от интенсивности света. Таким образом, колбочки двух типов вместе образуют прибор для измерения длины волны.
Почему же тогда двух типов рецепторов мало, чтобы полностью объяснить свойства нашего цветового зрения? Двух и в самом деле было бы достаточно, если бы мы имели дело только с монохроматическим светом — если бы мы согласились отказаться от таких вещей, как способность отличать цветной свет от белого. Наше зрение таково, что никакой монохроматический свет с любой длиной волны не выглядит белым. Это было бы невозможно при колбочках только двух типов. В случае красных и зеленых колбочек, продвигаясь от коротких к длинным волнам, мы постепенно переходим от стимуляции только зеленых к стимуляции только красных рецепторов со всеми промежуточными соотношениями между реакциями тех и других. Белый свет, состоящий, по существу, из смеси всех волн, должен в определенной мере стимулировать и красные, и зеленые колбочки. Таким образом, если монохроматический свет будет иметь длину волны, дающую то же соотношение реакций, то он будет неотличим от белого. Именно так обстоит дело при наиболее распространенной форме цветовой слепоты, когда человек имеет только два вида колбочек: независимо от того, какой из трех пигментов отсутствует, всегда найдется свет с какой-то длиной волны, неотличимый от белого. (Эти люди обладают дефектами цветового восприятия, но, конечно, не являются полностью цветнослепыми.)
Чтобы иметь цветовое зрение, подобное нашему, необходимо и достаточно иметь колбочки трех типов. Вывод о том, что у нас действительно именно три типа колбочек, был впервые сделан при исследовании особенностей цветового зрения человека, в результате ряда дедуктивных умозаключений, делающих честь человеческому интеллекту.
Теперь мы можем лучше понять, почему палочки не участвуют в восприятии цвета. При промежуточных уровнях освещенности могут функционировать как палочки, так и колбочки, но нервная система (если не считать редких искусственных ситуаций), по-видимому, не занимается вычитанием палочковых влияний из колбочковых. Колбочки сравнивают друг с другом, а палочки работают сами по себе. Если вы хотите убедиться в том, что палочки не передают информацию о цвете, проснитесь лунной ночью и оглядитесь. Хотя форму предметов вы сможете видеть довольно хорошо, цвета будут полностью отсутствовать. Удивительно, как мало людей осознают, что при слабом свете они обходятся без цветового зрения.
Увидим ли мы данный объект белым или цветным, определяется главным образом (но не всецело) тем, какие из трех типов колбочек активируются. Цвет — это результат неодинаковой стимуляции колбочек разного типа. Понятно, что свет с широкой спектральной кривой, например от солнца или от свечи, будет стимулировать колбочки всех трех типов (возможно, почти одинаково), и тогда ощущение окажется лишенным цвета, или «белым». Если бы нам удалось стимулировать колбочки только одного вида (что с помощью света сделать нелегко из-за перекрывания кривых поглощения), то результатом, как уже говорилось, был бы яркий цвет — фиолетовый, зеленый или красный в зависимости от вида стимулируемых колбочек. То, что максимум чувствительности тех колбочек, которые мы называем «красными», соответствует длине волны света, видимого нами как зеленовато-желтый (560 нм), связано, по-видимому, с тем, что такой свет возбуждает как зеленые, так и красные колбочки — из-за перекрывания кривых их спектральной чувствительности. Используя свет с большей длиной волны, мы можем более эффективно стимулировать красные колбочки по сравнению с зелеными.
Графики на рис. 120 резюмируют цветовые ощущения, возникающие при активации разных сочетаний колбочек светом различного спектрального состава. Первый и два последних примера должны убедительно показать, что ощущение «белого» цвета — результат примерно одинаковой стимуляции колбочек всех трех типов — может быть вызвано многими различными способами: и воздействием широкополосного света, и с помощью смеси узких спектральных полос, например желтого света с синим или красного с сине-зеленым. Два световых луча называют дополнительными, если их волновой состав и интенсивность подобраны так, что при смешении они дают ощущение «белого». В двух последних примерах синий и желтый, так же как красный с длиной волны 640 нм и сине-зеленый цвета являются дополнительными.
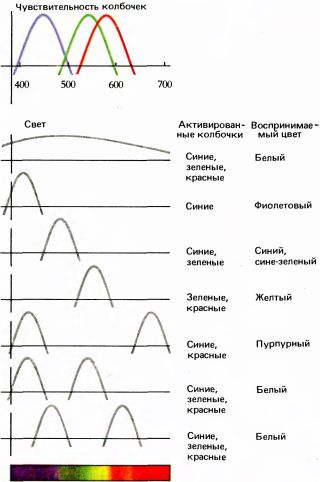
Рис. 120. Верхний график — «чувствительность колбочек» — повторяет график, приведенный на рис. 119. Далее указано, какие колбочки будут активироваться различными смесями окрашенного света и какие будут возникать ощущения.
Теории цветового зрения
Все сказанное выше о зависимости видимого цвета от стимуляции тех или иных колбочек основано на исследованиях, начатых Ньютоном в 1704 году и продолжающихся до сих пор. Изобретательность, которую проявил Ньютон в своих экспериментах, трудно переоценить: в работе, посвященной цвету, он при помощи призмы расщеплял белый свет; воссоединял его компоненты второй призмой, вновь получая белый свет; изготовил волчок с цветовыми секторами, при вращении которого опять-таки получался белый цвет. Эти открытия привели к осознанию того, что обычный свет состоит из непрерывного ряда лучей с различными длинами волн.
В XVIII столетии постепенно выяснилось, что всякий цвет можно получить путем смешения трех цветных компонентов в надлежащих пропорциях при условии, что длины их волн достаточно отличаются друг от друга. Представление о том, что любой цвет может быть «составлен» путем манипулирования тремя управляющими факторами (в данном случае путем изменения интенсивности трех различных лучей) получило название трихроматичности. В 1802 году Томас Юнг выдвинул четкую и простую теорию, объясняющую трихроматичность: он предположил, что в каждой точке сетчатки должны существовать по меньшей мере три «частицы» — крошечные структуры, чувствительные соответственно к красному, зеленому и фиолетовому. Длительный временной интервал между Ньютоном и Юнгом трудно объясним, но различные «дорожные препятствия» вроде, например, того факта, что желтая и синяя краски, смешиваясь, дают зеленую, не способствовали, конечно, ясности мышления. Решающие эксперименты, прямо и недвусмысленно подтвердившие, наконец, идею Юнга о том, что цвет должен определяться мозаикой трех видов детекторов в сетчатке, были проведены в 1959 году: Джордж Уолд и Пол Браун в Гарварде и Эдвард Мак-Никол и Уильям Маркс в Университете Джонса Гопкинса изучали под микроскопом способность отдельных колбочек поглощать свет с различной длиной волны и обнаружили три и только три типа колбочек. До этого ученые прилагали все усилия, используя менее прямые методы, и за несколько столетий фактически пришли к такому же результату, доказав теорию Юнга о необходимости именно трех типов колбочек и оценив их спектральную чувствительность. Применялись в основном психофизические методы: ученые выясняли, какие цветовые ощущения вызывают различные смеси монохроматических лучей, как влияет на цветовое зрение избирательное обесцвечивание рецепторов под действием монохроматического света, а также исследовали цветовую слепоту.
Изучение эффектов смешения цветов необычайно интересно — настолько его результаты удивительны и противоречат интуиции. Никто без предварительного знания не угадал бы разнообразные явления, иллюстрируемые на рис. 120 и 121, — например, не мог бы предсказать, что два пятна, ярко-синее и ярко-желтое, при наложении друг на друга сольются в белый цвет, неотличимый на глаз от цвета мела, или что зеленый и красный спектральные цвета при их объединении дадут желтый, почти неотличимый от монохроматического желтого цвета.

Рис. 121. С помощью трех диапроекторов и трех фильтров на экран проецируются три перекрывающихся пятна (красное, зеленое и синее). Красное и зеленое при наложении дают желтый цвет, синее и зеленое — бирюзовый, красное и синее — пурпурный, а все три вместе — белый цвет.
Прежде чем обсуждать другие теории цвета, нужно сообщить ряд дополнительных сведений о разнообразии цветов, которое эти теории призваны объяснить. Какие существуют цвета помимо цветов радуги? По моему мнению, имеются три вида таких цветов. Один вид — пурпурные, которые отсутствуют в радуге, но появляются при одновременной стимуляции красных и синих колбочек, т.е. при смешении длинно- и коротковолнового, или, грубо говоря, красного и синего света. Если к смеси спектрального красного и спектрального синего света — к пурпурному — мы добавим надлежащее количество зеленого, то мы получим белый цвет; поэтому мы говорим, что зеленый и пурпурный являются дополнительными. Можно, если угодно, представить себе круговую шкалу, включающую все цвета спектра от красного через желтый и зеленый до синего и фиолетового с последующим переходом к пурпурным цветам — сначала к синевато-пурпурному, затем к красновато-пурпурному и наконец опять к красному. Можно расположить эти оттенки так, чтобы дополнительные цвета располагались друг против друга. Понятие основных цветов не вписывается в эту схему: если определить основные цвета в соответствии с тремя типами рецепторов, то мы выделим зеленовато-желтый, зеленый и фиолетовый, т.е. оттенки, вряд ли согласующиеся с представлением о трех чистых базовых цветах. Но если под основными подразумевать три цвета, из которых можно получить любой другой оттенок, то упомянутые три цвета этому критерию удовлетворяют, как, впрочем, и любые другие три достаточно далеко отстоящие друг от друга цвета. Таким образом, ничто из сказанного выше не обосновывает представление о трех единственных основных цветах.
Второй тип цвета получается от добавления белого к любому цвету спектра или к пурпурному; мы говорим, что такое добавление «разбавляет» цвет, делает его бледнее — на профессиональном языке говорят, что белый уменьшает насыщенность цвета. Для подбора двух идентичных цветов мы должны сделать их одинаковыми по тону и насыщенности (выбрав, например, соответствующее положение на цветовом круге и затем добавив нужное количество белого), а потом уравнять по интенсивности. Таким образом, мы можем определить некоторый цвет, задав длину волны света (или в случае пурпурного — дополнительного к нему цвета), относительное содержание белого света и число, характеризующее интенсивность. Математически эквивалентная возможность определения цвета состоит в задании трех чисел, представляющих относительные влияния света на три типа колбочек. В любом случае необходимы три числа.
Типичным примером цвета третьего типа, не укладывающегося в приведенные выше объяснения, является коричневый. Я вернусь к нему позднее.
Герман Гельмгольц принял и отстаивал теорию Юнга, которая приобрела известность как теория Юнга—Гельмгольца. Между прочим, именно Гельмгольц объяснил, наконец, феномен, упомянутый в начале этой главы и заключающийся в том, что смесь желтой и синей красок дает зеленую. Вы можете легко убедиться, насколько это отличается от смешения желтого и синего света, проделав следующий опыт, для которого вам понадобятся всего лишь два диапроектора и немного желтого и синего целлофана. Сначала прикрепите желтый целлофан к линзе одного проектора, а синий — к линзе другого и наложите проецируемые изображения друг на друга. Отрегулировав относительные интенсивности, вы получите в зоне перекрывания чистый белый свет. Этот вид смешения цветов мы уже рассматривали; как мы тогда объяснили, белый свет возникает из-за того, что совместное воздействие желтого и синего света активирует все три системы колбочек с той же относительной эффективностью, что и широкополосный, или белый, свет. Теперь выключите один проектор и расположите оба фильтра перед другим; вы получите зеленый цвет. Чтобы понять, отчего так происходит, мы должны знать, что синий целлофан поглощает из белого света длинноволновую часть, т.е. желтый и красный, а остальную, которая выглядит синей, пропускает, в то время как желтый фильтр поглощает в основном синюю часть, а остальную, кажущуюся желтой, пропускает. Схема на рис. 122 показывает спектральный состав света, пропускаемого каждым фильтром. Обратите внимание на то, что в обоих случаях пропускаемый свет далек от монохроматического. Желтый свет — это не узкополосный спектральный желтый, а смесь спектрального желтого с более короткими зелеными, более длинными оранжевыми и красными волнами. Аналогичным образом, синий — это спектральный синий с примесью зеленого и фиолетового. Почему же в таком случае мы видим только желтый или только синий цвет? Дело в том, что ощущение желтого — результат одинаковой стимуляции красных и зеленых колбочек без какого-либо воздействия на синие колбочки; такую стимуляцию можно осуществлять как спектральным желтым (монохроматическим светом с длиной волны 580 нм), так и более широким волновым «мазком», который обычно свойствен пигментам, — нужно лишь, чтобы ширина спектра не была чрезмерно велика и спектр не содержал коротких волн, стимулирующих синие колбочки. Аналогичным образом, спектральный синий свет оказывает приблизительно такое же воздействие, как синий плюс зеленый плюс фиолетовый. Теперь при использовании двух фильтров, расположенных один перед другим, мы получим то, что? пропускают оба фильтра, т.е. зеленые лучи. Именно в этой области перекрываются приведенные на рис. 128 графики для широкополосного синего и желтого света. То же самое происходит с красками: желтая и синяя краски вместе поглощают весь свет, кроме зеленых участков, которые отражаются. Отметим, что если бы мы использовали в нашем опыте монохроматические желтый и синий фильтры, расположив их один перед другим, они не пропустили бы ничего. Смешивание происходит только потому, что свет, пропускаемый или отражаемый красящими веществами, имеет широкополосный спектральный состав.
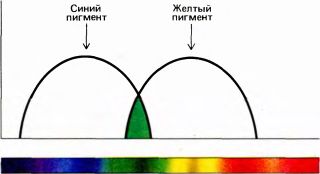
Рис. 122. Синий фильтр пропускает довольно широкую спектральную полосу с центром в области 480 нм, а желтый — такую же полосу с центром в области 580 нм. Оба фильтра вместе пропускают только общие для них лучи — свет в достаточно узкой полосе с центром около 530 нм, что дает зеленый цвет.
Резюмируем это многословное объяснение того, почему «желтый плюс синий равен зеленому», следующим кратким утверждением о цвете и красителях: два фильтра, расположенные один перед другим, или две смешанные краски совместно поглощают из белого света все, кроме волн средней длины, т.е. зеленого цвета.
Почему я обсуждаю здесь этот феномен? Отчасти потому, что это объясняет драматичный и сенсационный результат смешения желтого и синего с получением зеленого, но еще больше из-за исторической важности этого результата для подтверждения нашего понимания цветового зрения. Феномен этот физический; он связан с цветовым зрением и биологией примерно так же, как с ними связаны перекрещивание поляроидов и получение черного или добавление к кислоте синего лакмуса и получение красного, — говоря коротко, никак. И все-таки мысль о связи смешения цветов с цветовым зрением еще продолжает смущать многих, и это обусловлено представлением о том, что красный, желтый и синий — основные цвета, а зеленый — нет. Если какой-либо набор цветов и можно признать основным, так это четыре цвета — красный, синий, желтый и зеленый. Как мы убедимся в разделе, посвященном теории Геринга, основание, с которым все четыре цвета могут претендовать на звание основных, мало связано с тремя типами колбочек и значительно больше — с последующей переработкой информации в сетчатке и в мозгу.
(Этим нисколько не обесценивается знание живописцем того факта, что всего тремя красками можно имитировать большинство цветовых оттенков. Но даже мастер в своей области может ошибиться. В одной книге, посвященной ткацкому делу, в главе, излагающей теорию цвета, я нашел утверждение, что если вы смешаете в ткани желтые и синие нити, то получите зеленый цвет. На самом же деле получится серый цвет — по биологическим причинам.)
Цветовая слепота
Из работ Дж. Уолда, У. Раштона и многих других мы знаем, что в основе обычных форм цветовой слепоты, имеющейся примерно у 8 процентов мужчин, лежит отсутствие или нехватка одного или нескольких типов колбочек. Число возможных комбинаций отсутствия или количественного недостатка тех или иных колбочек делает цветовую слепоту весьма сложным объектом исследования.
Иногда цветовая слепота возникает в левом или правом поле зрения после локального инсульта в контралатеральном или ипсилатеральном полушарии. При этом, вероятно, повреждается какая-то высшая корковая зрительная зона, расположенная выше стриарной коры и зоны 18, — зона, названная V4 Семиром Зеки из Университетского колледжа. Об этих высших зонах мы поговорим в главе 10.
Теория Геринга
Параллельно теории цвета Юнга—Гельмгольца возникла и до недавнего времени казалась с ней несовместимой вторая научная школа. Эвальд Геринг (1834–1918) интерпретировал результаты смешения цветов, предположив, что в глазу и/или мозгу существуют три оппонентных процесса: один для ощущения красного и зеленого, другой для желтого и синего и третий, качественно отличный от двух первых, — для черного и белого. Геринга поразило отсутствие (их невозможно даже представить себе!) цветов, которые можно было бы описать как желтовато-синий или красновато-зеленый, а также «взаимное уничтожение» синего и желтого или красного и зеленого при их смешении в надлежащих пропорциях — цвет при этом полностью исчезает, т.е. возникает ощущение белого цвета. Геринг рассматривал красно-зеленый и желто-синий процессы как независимые в том смысле, что смесь синего и красного дает синевато-красный, или пурпурный; аналогично, смесь красного и желтого дает оранжевый, смесь зеленого и синего — синевато-зеленый, а смесь зеленого и желтого — зеленовато-желтый. В системе Геринга желтый, синий, красный и зеленый могут считаться «основными» цветами. Смотря на оранжевое, каждый может представить его себе как результат смешения красного и желтого, но никому не удается, глядя на красное или синее, видеть в нем результат смешения каких-либо других цветов. (Возникающее у некоторых людей ощущение, будто зеленый выглядит как желтый с добавкой синего, обусловлено, вероятно, их детским жизненным опытом, связанным с наборами красок.) Многим казалось, что представления Геринга о сине-зеленом и желто-синем процессах опираются всего лишь на интуитивные впечатления о цвете. Но поражает, насколько хорошо согласуются мнения людей, которых просили указать точку спектра, где представлен чистый синий цвет без какой-либо кажущейся примеси зеленого или желтого. То же самое можно сказать о желтом и зеленом цветах. Что касается красного цвета, то оценка испытуемых вновь совпадает, но в этом случае они настаивают на том, что нужно добавить немного фиолетового, чтобы устранить чуть заметную желтизну длинноволнового света. [Именно такой субъективный красный при добавлении к зеленому дает белый; обычный (спектральный) красный, добавленный к зеленому, дает желтый.]
Мы можем сравнить желто-синий и красно-зеленый процессы Геринга с двумя приборами вроде старинных вольтметров, стрелка одного из которых отклоняется влево при регистрации желтого и вправо при регистрации синего, а другой прибор ведет себя точно так же по отношению к паре «красный-зеленый». Цвет объекта при этом можно описать показаниями двух приборов. Третий антагонистический процесс Геринга (его можно представить себе как третий вольтметр) регистрирует соотношение черного и белого. Геринг понимал, что ощущение черного и серого порождается не просто отсутствием света, поступающего от некоторого объекта или поверхности, а возникает тогда и только тогда, когда от объекта приходит меньше света, чем в среднем от окружающих областей. Ощущение белого возникает только в том случае, если фон темнее и отсутствует цвет (это я уже обсуждал в главе 3 на примере выключенного телевизора). По теории Геринга черно-белый процесс предполагает пространственное сравнение или вычитание отражающих способностей, в то время как его желто-синий и красно-зеленый процессы происходят в одном определенном участке поля зрения и не связаны с окружением. (Геринг, несомненно, знал о взаимодействии соседних цветов, но его теория цвета, как она сформулирована в его поздних работах, не включает эти явления.) Мы уже видели, что черное и белое действительно представлены в сетчатке и в мозгу пространственно разделенными процессами возбуждения и торможения (on—off), которые в буквальном смысле слова антагонистичны.
Теория Геринга позволила объяснить не только все спектральные цвета и уровни насыщенности, но и такие цвета, как коричневый и оливково-зеленый, которые отсутствуют в радуге и даже не могут быть воспроизведены ни в одной из классических психофизических процедур смешения цветов, в которых мы с помощью диапроектора отбрасываем световые пятна на темный экран. Мы получим коричневый цвет лишь в том случае, если желтое или оранжевое световое пятно будет окружено в среднем более ярким светом. Возьмите любую коричневую поверхность, посмотрите на нее через свернутый трубкой кусок черной бумаги, чтобы исключить все ее окружение, и вы увидите желтый или оранжевый цвет. Мы можем считать коричневый цвет смесью черного, получаемого только в условиях пространственного контраста, с оранжевым или желтым. По терминологии Геринга, при этом работают по меньшей мере две системы — черно-белая и желто-синяя.
Теория Геринга о трех оппонентных системах — красно-зеленой, желто-синей и черно-белой — в его время и еще полстолетия рассматривали как альтернативную по отношению к трехкомпонентной («красный, зеленый, синий») теории Юнга — Гельмгольца. Сторонники каждой из них были, как правило, весьма фанатичны и зачастую чрезмерно эмоциональны. Физики обычно примыкали к лагерю Юнга — Гельмгольца, быть может потому, что их привлекали количественные аргументы (такие, например, как системы линейных уравнений) и отталкивали доводы, связанные с чистотой цветов. Психологи часто были на стороне Геринга, вероятно в связи с тем, что им приходилось иметь дело с более широким разнообразием психофизических феноменов. Теория Геринга, казалось, содержала доводы в пользу либо четырех типов рецепторов (красный, зеленый, желтый и синий), либо трех (черно-белый, желто-синий и красно-зеленый); оба варианта противоречили накапливавшимся данным, которые подкрепляли исходную гипотезу Юнга. Ретроспективно можно сказать, как отмечают современные психофизики Лео Гурвич и Доротея Джеймсон, что одна из трудностей была связана с отсутствием до 1950-х годов каких-либо прямых физиологических данных о тормозных механизмах в сенсорных системах. Такие данные появились лишь тогда, когда стала возможной регистрация активности одиночных нейронов.
Представив себе вольтметры, измеряющие положительные величины отклонением вправо, а отрицательные — влево, вы сможете понять, почему теория Геринга предполагает наличие тормозных механизмов. Желтый и синий цвета взаимно антагонистичны; смешиваясь, они уничтожают друг друга, а если стрелка красно-зеленой системы тоже указывает на нуль, то цвет отсутствует. Геринг в известном смысле на пятьдесят лет опередил свое время. Как это случалось и раньше в истории науки, две теории, на протяжении десятилетий казавшиеся несовместимыми, обе оказались верны. В конце прошлого столетия никто не мог предположить, что представления Юнга — Гельмгольца окажутся верными для рецепторного уровня, а идеи Геринга об оппонентных процессах — для последующих уровней зрительной системы. Теперь стало ясно, что эти две формулировки не исключают друг друга: обе они предполагают наличие системы с тремя переменными — это три типа колбочек в теории Юнга—Гельмгольца и три измерительных прибора или процесса в теории Геринга. Что нас сегодня изумляет, так это то, что Геринг, опираясь на столь ограниченный фактический материал, сумел сформулировать теорию, так хорошо согласующуюся с нейронной организацией центральных механизмов цветового зрения. Тем не менее специалисты по цветовому зрению все еще разделены на два лагеря: одни считают Геринга пророком, а другие видят в упомянутом соответствии лишь случайную удачу. Я наверняка наживу врагов среди тех и других, так как занимаю нейтральную позицию и только чуть-чуть склоняюсь в пользу первого мнения.
Цвет и пространство
В главе 3 мы видели, что восприятие объекта как белого, черного или серого зависит от его относительной способности отражать свет по сравнению с другими предметами, находящимися в поле зрения. Таким образом, свойства широкополосных клеток нижних уровней зрительной системы — ганглиозных клеток сетчатки и клеток коленчатого тела — позволяют в значительной мере объяснить восприятие черного, белого и серого: именно такое сравнение они осуществляют при помощи своих рецептивных полей с центром и периферией. Несомненно, именно в этом состоит третий, пространственно оппонентный черно-белый процесс Геринга. То, что пространственная переменная важна и для восприятия других цветов, начали впервые осознавать еще столетие назад; однако аналитический подход к этому вопросу стал разрабатываться только в последние десятилетия, в основном усилиями таких психофизиков, как Лео Гурвич и Доротея Джеймсон, Дин Джадд и Эдвин Лэнд. Лэнд с его глубоким интересом к проблемам освещения и фотографии был, естественно, заинтригован неспособностью фотокамеры компенсировать различия в источниках света. Если фотопленка сбалансирована так, что изображение белой рубашки выглядит белым при свете вольфрамовых ламп накаливания, то та же рубашка под синим небом будет светло-голубой; если же пленка предназначена для естественного света, рубашка при обычном электрическом свете будет розовой. При изготовлении хорошей цветной фотографии мы должны учитывать не только интенсивность света, но и его спектральный состав — будет ли свет голубоватым или красноватым. Если мы знаем это, мы сможем установить выдержку и диафрагму с учетом интенсивности и подобрать пленку или фильтры с учетом цветового баланса. В отличие от фотокамеры наша зрительная система делает все это автоматически; она настолько хорошо решает эту задачу, что обычно мы даже не осознаем, что такая проблема существует. Белая рубашка выглядит белой, несмотря на большие сдвиги в спектральном составе света при переходе от освещения солнцем в зените к освещению заходящим солнцем, вольфрамовой или флуоресцентной лампой. То же постоянство сохраняется для окрашенных объектов, и этот феномен применительно к цветному и белому называют константностью цвета. Хотя константность известна уже давно, демонстрации Лэнда в 50-х годах явились большим сюрпризом даже для нейрофизиологов, физиков и большинства психологов.

Рис. 123. Во многих своих экспериментах Эдвин Лэнд использовал мозаику из цветных бумажных полосок в стиле Мондриана. Целью экспериментов было доказательство того, что воспринимаемые цвета сохраняют поразительное постоянство, несмотря на заметные изменения в соотношении красных, зеленых и синих лучей, использованных для освещения мозаики.
Каковы же эти демонстрации? В типичном эксперименте мозаику из прямоугольных кусков бумаги разного цвета, напоминающую рисунки Мондриана, освещают тремя диапроекторами, один из которых снабжен красным, другой зеленым и третий синим фильтрами. Каждый проектор имеет регулируемый источник света, так что интенсивность его можно изменять в широких пределах. В остальном комната должна быть полностью затемнена. Если все три проектора установить на среднюю интенсивность, то цвета будут выглядеть примерно так же, как при дневном свете. Удивительно, что точная установка как будто не имеет значения. Выберем зеленый участок мозаики и фотометром, точно измерим интенсивность приходящего от него света, когда включен только один проектор. Затем повторим измерение со вторым проектором, а потом с третьим. Это даст нам три числа, характеризующих свет, отражаемый при включении всех трех проекторов. Выберем теперь другой участок, например оранжевый, и отрегулируем поочередно интенсивность каждого проектора так, чтобы показания фотометра для оранжевого участка совпадали с теми, которые мы получили ранее для зеленого. Таким образом, при включении трех проекторов свет, приходящий теперь от оранжевого участка, идентичен по составу тому, который минутой ранее приходил от зеленого участка. Что мы ожидаем увидеть? Рассуждая примитивным образом, мы скажем, что оранжевый участок превратился в зеленый. Но он все-таки выглядит оранжевым — его цвет даже нисколько не изменился. Мы можем повторить этот эксперимент с любыми двумя участками. Вывод таков: не столь важно, на какую интенсивность света установлены три проектора, пока от каждого из них поступает какой-то свет. Почти в любом случае мы заметим лишь очень малые изменения воспринимаемого цвета.
Такие эксперименты убедительно показали, что ощущение, возникающее в какой-либо части поля зрения, зависит как от света, приходящего от этой части, так и от света, приходящего от остальных участков. Иначе как мог бы свет одинакового спектрального состава вызывать в одном случае ощущение зеленого, а в другом — ощущение оранжевого? Принцип, применимый к черному, белому и серому и столь четко сформулированный Герингом, оказывается верным и в отношении цвета. Для цвета мы имеем оппонентность не только локальную (красный/зеленый и желтый/синий), но также и пространственную: красный/зеленый в центре против красного/зеленого на периферии и аналогичную оппонентность для желтого/синего.
В 1985 году Дэвиду Инглу в лаборатории Лэнда удалось в аквариуме с подводной мозаикой из разноцветных прямоугольников обучить золотую рыбку подплывать к участку определенного цвета. Он обнаружил, что рыбка плывет к одному и тому же цвету, например синему, независимо от спектрального состава света: она, как и мы, выбирает синий участок, даже если свет от него идентичен по составу свету, который в предыдущей пробе при ином источнике света приходил от желтого лоскута, отвергаемого рыбкой. Таким образом, рыба тоже выбирает участок поверхности по его собственному цвету, а не по спектральному составу отражаемого им света. Это означает, что феномен константности цвета не следует считать каким-то усовершенствованием, недавно добавленным в ходе эволюции к цветоощущению некоторых высших млекопитающих, в том числе и человека; наличие его у рыб указывает на то, что это примитивный, весьма общий аспект цветового зрения. Очень заманчиво (и довольно несложно) было бы проверить, обладают ли той же способностью насекомые с цветовым зрением. Я думаю, что это именно так.
Лэнд и его группа (среди прочих Дж. Мак-Кэнн, Н. Доу, М. Бёрнс и X. Перри) разработали несколько процедур для предсказания видимого цвета некоторого объекта по спектрально-энергетическому составу света от всех точек поля зрения, но без какой-либо информации о источнике света. Расчет заключается в том, что для каждого из трех отдельных проекторов определяют отношение света, приходящего от места, цвет которого нужно предсказать, к усредненному свету, приходящему от окружения. (Площадь «окружения», которую нужно учитывать, в разных вариантах теории Лэнда различна. В новейшем варианте предполагается, что влияние окружающих участков уменьшается с расстоянием.) Получаемая тройка чисел — отношения берутся для каждого проектора — однозначно определяет цвет данного места. Любому цвету, таким образом, можно поставить в соответствие некоторую точку в трехмерном пространстве, координатными осями которого будут три отношения, полученные для красного, зеленого и синего света. Чтобы сделать формулировку как можно более реалистичной, три источника света подбираются в соответствии с кривыми спектральной чувствительности трех типов колбочек человека.

Рис. 124. В экспериментах Дэвида Ингла золотая рыбка обучалась подплывать за вознаграждение — кусочек печенки — к полоске заданного цвета. Она плывет, например, к желтой полоске независимо от соотношения интенсивностей света от трех проекторов. Такое поведение поразительно сходно с константностью восприятия цвета у человека.
То, что таким образом можно вычислить цвет, означает наличие константности цвета, поскольку для каждого проектора подсчитывается соотношение света от одной области к свету от усредненного окружения. Точная установка силы света в проекторах более не важна: единственным условием остается то, что мы должны иметь какой-то свет от каждого проектора — иначе нельзя будет вычислить никакого отношения. Одно из следствий всего этого заключается в том, что для появления цвета необходимы различия в спектральном составе света в пределах поля зрения. Для восприятия цвета нам нужны цветовые границы, точно так же как нужны границы освещенности для восприятия черного и белого. Вы можете легко убедиться в этом, снова использовав два диапроектора. Поставьте перед одним из проекторов красный фильтр (подойдет красный целлофан) и осветите любую группу предметов. Я предпочитаю брать белую или желтую рубашку и яркий красный галстук. При таком освещении ни рубашка, ни галстук не выглядят вполне красными: и то и другое кажется розоватым и как бы полинявшим. Теперь осветите ту же комбинацию предметов вторым проектором, прикрыв его синим целлофаном. Рубашка будет выглядеть бледно-голубоватой, а галстук — черным: красные объекты не отражают коротких световых волн. Вернитесь к красному проектору и убедитесь еще раз, что галстук не выглядит особенно красным. Теперь добавьте синий проектор. Вы знаете, что при добавлении синего света вы не получите от галстука какого-либо дополнительного отражения — вы только что это продемонстрировали, — но при включении синего проектора красный галстук внезапно засверкает хорошим ярким красным цветом. Это убедит вас в том, что галстук делают красным не только лучи, приходящие от него самого.
Эксперименты со стабилизированными цветовыми границами согласуются с представлением о том, что вообще для видения цвета необходимы различия на границах. Альфред Ярбус, имя которого упоминалось в главе 4 в связи с движением глаз, в 1962 году показал, что если смотреть на синее пятно, окруженное красным фоном, стабилизация границы пятна на сетчатке приводит к его исчезновению: синее пропадает, и все, что можно видеть, — это красный фон. Стабилизация границ на сетчатке явно делает их неэффективными, а без них исчезает и цвет.
Эти психофизические доказательства того, что для восприятия цвета нужны различия в спектральном составе света от разных участков поля зрения, указывают на возможное наличие в наших сетчатках или мозгу клеток, чувствительных к цветовым границам. Этот довод аналогичен тому, который приводился нами в главе 4 по поводу восприятия черных или белых объектов. Если на каком-то уровне нашей зрительной системы информация о цвете передается только на линиях цветового контраста, то клетки с рецептивными полями, лежащими целиком внутри областей однородного цвета, будут бездействовать. Результат — экономия при обработке информации. Таким образом, от передачи сведений о цвете лишь на границах мы получаем два преимущества: во-первых, цвет не меняется при изменениях освещения, так что мы узнаём о свойствах рассматриваемых объектов без искажений, вносимых источником света; во-вторых, информация обрабатывается экономным способом. Теперь мы можем задаться вопросом: почему система эволюционировала именно таким образом? Была ли главным фактором эволюции необходимость константности цвета, а экономия — лишь сопутствующей выгодой? Или, наоборот, экономия играла ведущую роль, а константность — второстепенную. Второе предположение многим может показаться более убедительным: вряд ли эволюция могла предвидеть появление ламп накаливания или флуоресцентного освещения, да и наши рубашки вовсе не были такими белыми, пока не появились современные моющие средства.
Физиология цветового зрения: ранние результаты
Первая физиологическая информация на клеточном уровне была получена спустя 250 лет после Ньютона в исследованиях шведско-финско-венесуэльского физиолога Гуннара Светихина, который в 1956 году на костистой рыбе осуществил внутриклеточную регистрацию активности нейронов сетчатки — сначала он принял их за колбочки, но они оказались горизонтальными клетками. На освещение сетчатки эти клетки отвечали только медленными потенциалами (потенциалов действия не наблюдалось). Как показано на рис. 125, Светихин обнаружил три типа клеток: первый тип, названный им L-клетками, гиперполяризовался при световой стимуляции независимо от спектрального состава света; второй тип, названной r-g-клетками, гиперполяризовался волнами малой длины с максимумом ответа на зеленый свет и деполяризовался волнами большой длины с максимумом ответа на красный свет; третий тип, названный с учетом теории Геринга y-клетками, отвечал по типу клеток r-g, но с максимумом гиперполяризации на синий и максимумом деполяризации на желтый свет. У клеток r-g и y-b белый свет вызывал лишь слабые и быстро затухающие ответы, как и следовало ожидать ввиду широкополосного спектрально-энергетического состава белого света. Кроме того, у клеток обоих этих типов, которые мы можем назвать оппонентно-цветовыми клетками, свет с некоторой промежуточной длиной волны, называющейся точкой перекреста, не вызывал никакой реакции. Поскольку эти клетки реагируют на окрашенный, но не на белый свет, они, вероятно, связаны с цветовыми ощущениями.
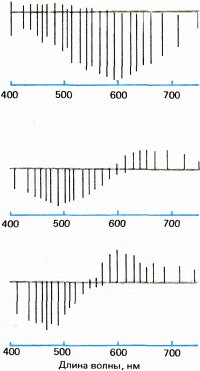
Рис. 125. Гуннар Светихин и Эдуард Мак-Никол регистрировали реакции горизонтальных клеток на цвет у костистой рыбы. Отклонения вниз от серой линии соответствуют гиперполяризации, а отклонения вверх — деполяризации.
В 1958 году Рассел де Валуа и его сотрудники зарегистрировали ответы, поразительно сходные со светихинскими, от клеток наружного коленчатого тела макака. Ранее с помощью поведенческих тестов де Валуа показал, что цветовое зрение макака и человека почти одинаково; например, соотношение, в котором нужно смешать два цветных луча, чтобы получить третий цвет, почти идентично у обоих видов. Поэтому можно думать, что макаки и люди обладают сходными механизмами на нижних уровнях зрительной системы, и мы, по-видимому, вправе сравнивать психофизику цвета у человека с физиологией у макака. Де Валуа обнаружил, что многие клетки коленчатого тела активировались рассеянным монохроматическим светом в диапазоне от одного конца спектра до точки перекреста, где реакция отсутствовала, и подавлялись светом во втором диапазоне — от точки перекреста до другого конца спектра. И вновь аналогия с цветовыми процессами Геринга была полной: де Валуа выявил оппонентно-цветовые клетки двух типов, красно-зеленые и желто-синие; у каждого типа смешение двух световых волн, длины которых на волновой шкале располагались симметрично относительно точки перекреста, приводило к взаимному погашению реакций точно так же, как в восприятии добавление синего к желтому или зеленого к красному порождает белый. Результаты де Валуа особенно хорошо согласовались с формулировками Геринга, поскольку две группы цветовых клеток имели максимумы реакции и точки перекреста точно в тех местах спектральной шкалы, чтобы одна группа могла отражать «желто-синие» свойства падающего света, а другая группа — «красно-зеленые».
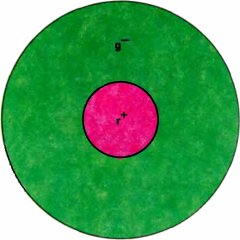
Рис. 126. В типичном рецептивном поле типа 1 центр посылает возбуждающие сигналы от красных колбочек, а периферия — тормозящие сигналы от зеленых колбочек.
Следующий шаг состоял в том, чтобы рассмотреть рецептивные поля этих клеток, используя вместо рассеянного света маленькие цветные пятнышки; это мы и сделали в 1966 году совместно с Торстеном Визелом. У рецептивных полей большинства оппонентно-цветовых клеток де Валуа обнаружилась удивительная организация, которая до сих пор ставит нас в тупик. Клетки, как и у кошек по данным Куффлера, имели поля, разделенные на два антагонистических участка — центр и периферию; центр мог быть типа on или off. В типичном случае центр поля представлен исключительно красными, а тормозящая периферия — исключительно зелеными колбочками. Поэтому при красном свете как маленькое, так и большое пятно вызывает энергичную реакцию, поскольку центр избирательно чувствителен к длинноволновому свету, а периферия на него почти не реагирует; при коротковолновом свете маленькие пятна дают лишь очень слабую реакцию или не вызывают ее вовсе, а большие порождают сильное торможение с off-реакциями. При белом свете, содержащем короткие и длинные волны, малые пятна вызывают on-реакции, а большие не вызывают ответа.
Хотя нашим первым впечатлением было, что такая клетка из центра должна получать входные сигналы от красных, а с периферии — от зеленых колбочек, теперь кажется вероятным, что полное рецептивное поле включает две перекрывающиеся системы, как это показано на рис. 127. Как красные, так и зеленые колбочки распределены по довольно широкой круговой области, причем их количество максимально в центре и убывает по мере удаления от него. В центре красные колбочки сильно преобладают, но к периферии число их падает значительно быстрее числа зеленых колбочек. Поэтому небольшое пятнышко, вспыхивающее в центре и содержащее длинноволновую компоненту, будет весьма мощным стимулом для красной системы; даже если оно стимулирует и зеленые колбочки, их число по сравнению с общим числом связанных с клеткой зеленых колбочек будет слишком мало, чтобы составить какую-либо конкуренцию красной системе. Те же соображения приложимы и к клеткам с центром и периферией, описанным в главе 3, рецептивные поля которых тоже должны состоять из двух оппонентных круговых перекрывающихся зон с разными формами кривых зависимости чувствительности от координаты. Таким образом, периферия, вероятно, имеет не кольцеобразную, как думали вначале, а круговую форму. В отношении этих оппонентно-цветовых клеток обезьян предполагается (правда, без достаточных данных), что периферические области отражают вклад горизонтальных клеток.
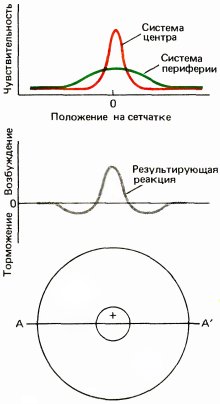
Рис. 127. Графики вверху отражают зависимость чувствительности нейрона (измеренной, например, по реакции на неподвижное очень маленькое световое пятнышко) от положения стимула на сетчатке вдоль линии AA, проходящей через центр рецептивного поля. Для клетки с r-центром и g–-периферией красное пятнышко дает островершинную кривую, а зеленое — значительно более широкую кривую. Нижний график отображает реакцию на белый или желтый свет, стимулирующий обе оппонентные системы, так что происходит вычитание вкладов двух систем. При этом красные колбочки доминируют в центре, что приводит здесь к on-реакциям, а зеленые — на периферии, что приводит к off-реакциям.
Реакция на диффузный свет — в данном случае on-реакции на красный, off-реакции на синий и зеленый и отсутствие реакции на белый свет — ясно показывают, что такая клетка должна регистрировать информацию о цвете. Но реакции на определенного рода белые края и отсутствие реакций на диффузный свет убеждают в том, что эта клетка связана также и с восприятием формы. Мы называем эти цветооппонентные клетки с центром и периферией клетками «типа 1».
Наружное коленчатое тело обезьяны, если вспомнить сведения из главы 4, содержит шесть слоев, причем четыре верхних слоя содержат множество мелких клеток, а два нижних — меньшее число крупных клеток. Мы находим клетки описанного выше типа 1 в верхних, или мелкоклеточных, слоях. Они различаются по типу колбочек, входящих в их центральную и периферийную системы, и по характеру центра, который может быть возбуждающим или тормозящим. Пример, представленный на рис. 126, мы можем обозначить как r+g– (с красным центром и зеленой периферией). Среди подтипов клеток, получающих входы от красных и зеленых колбочек, мы находим все четыре возможных варианта: r+ g–, r– g+, g+ r–, g– r+. Вторая группа клеток получает входы от синих колбочек из центра поля и от комбинации красных и зеленых (или, быть может, только от зеленых) — с периферии. Мы называем такие клетки «сине-желтыми», причем слово «желтые» здесь употреблено для краткости вместо «красные плюс зеленые».
В четырех верхних слоях мы находим клетки еще двух типов. Клетки типа 2 составляют около 10 процентов нейронной популяции и имеют рецептивные поля, состоящие только из центра. Повсюду в этом центре у некоторых клеток выявляется красно-зеленая, а у остальных — сине-желтая оппонентность. Рецептивные поля еще примерно у 15 процентов клеток в четырех верхних слоях коленчатого тела и у всех клеток в двух нижних (крупноклеточных) слоях имеют центр и периферию, но эти клетки не проявляют цветовых предпочтений; создается впечатление, что центр и периферия их полей получают одинаковый относительный вклад от всех трех видов колбочек. Мы считаем эти клетки широкополосными и в верхних слоях называем их клетками типа 3.
Все эти данные поразительно согласуются с моделью Геринга: кроме двух классов клеток с цветовой оппонентностью мы имеем также третий класс, вовсе не обладающий этим свойством, но с широкополосной пространственной оппонентностью. Что как будто не согласуется ни с какой теорией, так это пространственная организация полей у оппонентно-цветовых клеток — клеток типа 1. На первый взгляд можно подумать, что эта организация каким-то образом связана с цветовым контрастом, т.е. с тенденцией одного цвета, например синего, казаться более ярким в окружении другого цвета, например зеленого, или с тем, что кусок серой бумаги кажется желтоватым, если мы видим его на синем фоне. Но минутное размышление убедит вас, что клетки коленчатого тела едва ли могут быть полезны для этого вида цветового контраста: описанная выше клетка с r+-центром и g–-периферией не будет сильно возбуждена красным пятном на зеленом фоне — она ответит очень слабо или совсем не ответит, так как один эффект уничтожится другим; произойдет обратное тому, что нужно для усиления цветового контраста.
О клетках типа 1 можно лишь утверждать, что по своей численности они должны быть наиболее важным для мозга источником цветовой информации, хотя эта информация представлена каким-то странным образом. Их возможная работа совместно с клетками типа 3 очень хорошо согласуется с представлениями Геринга о двух оппонентных цветовых системах и одной оппонентной пространственной системы. Вероятно, они играют также важную роль в восприятии тонких деталей формы объектов, поскольку единственные другие клетки коленчатого тела с маленькими центрами полей — это широкополосные клетки типа 3, которых в десять раз меньше. Как мы видели в главе 6, остроту зрения, т.е. способность различать малые объекты, можно измерять несколькими способами; можно, например, определять наименьшее расстояние между двумя точками, при котором они еще не сливаются, или наименьший видимый разрыв в окружности (так называемом кольце Ландольта). Острота, измеренная по любому из этих способов, оказывается для центральной ямки порядка 0,5 угловой минуты, или около 1 миллиметра с расстояния 8 метров. Этот показатель хорошо согласуется с расстоянием между двумя колбочками в центральной ямке. Клетки коленчатого тела типа 1, получающие входные сигналы от зоны центральной ямки, имеют рецептивные поля с центрами порядка 2 угловых минут в диаметре. Кажется вероятным, что центр поля составляет здесь всего одна колбочка. Таким образом, мы находим разумное соответствие между остротой зрения и наименьшими размерами центра рецептивного поля у клеток наружного коленчатого тела.
Вентральная (нижняя) пара слоев коленчатого тела отличается от четырех дорсальных слоев тем, что состоит исключительно из клеток с широкополосными центрами полей. Эти клетки проявляют любопытную форму цветовой оппонентности, смысл которой никто не понимает и о которой я не буду говорить подробнее. Большинство считает эти клетки «слепыми» к цвету. Центры их полей в несколько раз крупнее, чем центры нейронов мелкоклеточных слоев, и у них есть ряд других интересных особенностей. В настоящее время мы подозреваем, что эти клетки обслуживают отделы мозга, играющие важную роль в восприятии формы, глубины и движения. Развитие этой темы увело бы нас далеко в сторону от цвета, и потребовалось бы написать еще одну книгу.
Большинство типов клеток, описанных для наружного коленчатого тела, встречается также и в сетчатке. В коленчатом теле они больше обособлены, что облегчает их изучение. Вклад коленчатого тела в анализ зрительной информации у обезьян до сих пор не известен.
Нейронная основа константности цвета
Поскольку клетки типа 1 наружного коленчатого тела как будто не приспособлены для цвето-пространственных сравнений, нам, по-видимому, следует выйти на более высокие уровни зрительной системы. Для проверки идеи, согласно которой такие сравнения могли бы происходить в коре головного мозга, группа Лэнда, Маргарет Ливингстон и я изучали человека, у которого была произведена перерезка мозолистого тела с целью лечения эпилепсии. Между областями, разделенными средней вертикалью поля зрения, пространственно-цветовые взаимодействия у него отсутствовали: на цвет пятна, расположенного чуть левее точки, на которую смотрел этот испытуемый, не влияли даже резкие цветовые изменения в правом зрительном поле, тогда как нормальные испытуемые сообщали в таких случаях о заметных изменениях. Это означает, что цвето-пространственные взаимодействия, по-видимому, не могут происходить в самой сетчатке. Хотя никто всерьез и не утверждал обратного, вопрос продолжал дискутироваться, и было приятно получить какие-то экспериментальные данные. Результаты этого эксперимента согласуются с тем фактом, что нам не удалось обнаружить в сетчатке такие ганглиозные клетки, которые вполне могли бы участвовать в цвето-пространственных взаимодействиях.
У золотой рыбки, осуществляющей пространственные сравнения не хуже нас с вами, коры головного мозга практически нет. Возможно, рыбка в отличие от нас выполняет эти сравнения с помощью своей сетчатки. Открытие в сетчатке рыб двойных оппонентных клеток (Н. Доу, 1968) как будто подтверждает это. У обезьян (см. следующий раздел) мы находим такие клетки в коре, но их нет ни в сетчатке, ни в наружном коленчатом теле.
Пузырьки
Примерно к 1978 году первичная зрительная кора обезьян с ее простыми и сложными клетками, с нейронами, реагирующими на концы линий, с глазодоминантными и ориентационными колонками представлялась достаточно изученной. Но неожиданной особенностью ее физиологии казалось то, что как будто лишь немногие клетки здесь небезразличны к цвету. Когда мы картировали рецептивное поле простой или сложной клетки, используя белый свет, а затем повторяли опыт с окрашенными пятнами или полосками, то результаты, как правило, были одинаковыми. Однако у некоторых клеток, составляющих, наверное, лишь десятую долю всех нейронов верхнего слоя коры, обнаруживались явные цветовые предпочтения — энергичные ответы на ориентированную полоску, скажем, красного цвета при полном отсутствии ответов на другие цвета или даже на белый цвет. Ориентационная избирательность этих клеток была не ниже, чем у клеток, нечувствительных к цвету, но большинство нейронов к цвету были безразличны. Все это казалось особенно странным ввиду того, что в наружном коленчатом теле очень большая доля клеток кодирует цвет, а коленчатое тело служит главным источником информации для зрительной коры. Трудно было понять, что могло происходить с этой цветовой информацией в коре.
Неожиданно в 1978 году все изменилось. Нейроанатом из Сиэтла Маргарет Уонг-Райли обнаружила, что при окрашивании коры на фермент цитохромоксидазу в верхних слоях выявляется никогда не наблюдавшаяся ранее неоднородность — периодические темноокрашенные участки шириной около четверти миллиметра, разделенные промежутками около полумиллиметра. Цитохромоксидазу — участвующий в метаболизме фермент — содержат все клетки, и никто не мог даже подумать, что гистохимическая реакция на этот фермент позволит увидеть в коре что-либо интересное. Когда Уонг-Райли прислала нам свои микрофотографии, Торстен Визел и я заподозрили, что мы видим колонки глазодоминантности в поперечном разрезе и что большинство монокулярных клеток по каким-то причинам метаболически более активны, чем бинокулярные. Мы положили снимки в ящик стола и постарались забыть о них.
Прошло несколько лет, прежде чем нам или кому-либо другому довелось изучать с помощью той же реакции срезы первичной зрительной коры, параллельные ее поверхности. Когда это приблизительно одновременно было, наконец, сделано двумя группами (Анитой Хендриксон и А. Хамфри в Сиэтле и Дж. Хортоном и мной в Бостоне), выявился — ко всеобщему полному изумлению — узор, напоминающий материал в горошек. Пример показан на рис. 129. Вместо полосок мы увидели похожие на пузырьки образования, которые ни с чем из известного ранее не связывались. Неоднородности Уонг-Райли называли всеми мыслимыми именами: точки, пуфы, мушки, пятнышки. Мы назвали их «пузырьками» [blobs — это слово наглядно, узаконено (оно есть даже в оксфордском словаре английского языка) и как будто раздражает наших конкурентов].

Рис. 128. На этом поперечном срезе стриарной коры видны слои, окрашенные для выявления фермента цитохромоксидазы. Более темные области в слоях 2 и 3 (вертикальные полоски вверху) — «пузырьки».
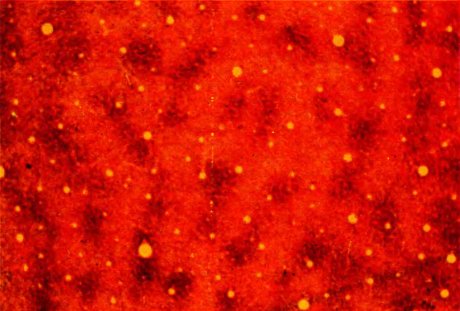
Рис. 129. Темные пятна представляют собой пузырьки, видимые «в плане»; около 50 таких пузырьков образуют характерный узор. Срез сделан через слой 3 параллельно поверхности коры на глубине около 0,5 мм. Срез проходит по границе между полями 17 (левая и средняя части) и 18 (правая часть), где пузырьков нет. (Желтые кружки — поперечные сечения кровеносных сосудов.)
Следующая задача была очевидной: мы должны были снова регистрировать ответы клеток в стриарной коре, гистологически контролируя эксперимент окраской на цитохромоксидазу, и попытаться выявить нечто особенное у клеток, находящихся в пузырьках. В 1981 году Маргарет Ливингстон и я принялись за эту работу. Результат оказался совершенно неожиданным. Проходя расстояние в четверть миллиметра, равное диаметру пузырька, можно исследовать примерно пять или шесть клеток. Всякий раз, когда мы пересекали пузырек, попадавшиеся на пути электрода клетки были полностью лишены ориентационной избирательности, что заметно контрастировало с высокой ориентационной избирательностью клеток, расположенных вне пузырьков.
Этому отсутствию ориентационной специфичности можно было бы дать два объяснения. Во-первых, эти клетки могли неизбирательно получать входные сигналы от соседних ориентационных клеток, лежащих вне пузырьков, и поэтому еще способны были отвечать специфической реакцией на линии (полоски и т.п.), но после объединения всех возможных ориентаций всякое предпочтение какой-либо из них исчезало. Во-вторых, они могли быть похожими на клетки коленчатого тела или на корковые клетки слоя 4C и, таким образом, быть проще внепузырьковых ориентационно-избирательных клеток. Вскоре вопрос был разрешен: оказалось, что большинство этих клеток имеет рецептивные поля с центром и периферией. Несколько дополнительных экспериментов убедили нас в том, что многие из них участвуют в кодировании цвета.
Больше половины пузырьковых клеток имели оппонентно-цветовые рецептивные поля с центром и периферией, но вели себя явно более сложным образом, чем клетки типа 1 наружного коленчатого тела. Они практически не отвечали на белые пятна любых размеров и формы. Но на небольшие цветные пятна, вспыхивающие в центре рецептивного поля, они энергично реагировали в одном диапазоне длин волны и затормаживались в другом диапазоне; некоторые активировались длинными волнами (красным светом) и подавлялись короткими (зеленым и синим светом), другие вели себя противоположным образом. Как и среди клеток коленчатого тела, мы могли, в зависимости от положения максимальных реакций на спектральной шкале, выделить два класса — красно-зеленые и сине-желтые клетки. (Здесь, как и ранее, слова «красный», «зеленый» и «синий» указывают на соответствующие типы колбочек, а слово «желтый» — на параллельные входы от красных и зеленых колбочек.) Таким образом, эти клетки весьма напоминали оппонентно-цветовые клетки коленчатого тела, обладающие только центром (тип 2). Но в отличие от клеток типа 2 эти цветокодирующие пузырьковые клетки чаще всего не отвечали на большие белые или цветные пятна, каков бы ни был спектральный состав света. Они вели себя так, как если бы центральная система рецептивного поля была у них окружена кольцом оппонентности. Если говорить о наиболее распространенном типе клеток, то центр типа r+g–, казалось, был окружен у них кольцом типа r–g+.
Мы с Маргарет Ливингстон назвали эти клетки двойными оппонентными из-за их красно-зеленой или желто-синей оппонентности в центре и антагонизма периферии по отношению к любой реакции в центре, будь она типа on или off. Поэтому они не отвечают не только на белый свет в любой геометрической конфигурации, но и на большие пятна, независимо от их спектрального состава. Центры рецептивных полей, как и у клеток типа 2, были в несколько раз больше, чем у клеток коленчатого тела типа 1. Как уже упоминалось, Н. Доу ввел термин двойные оппонентные для клеток, обнаруженных им в сетчатке золотой рыбки. Он предполагал, что сходные клетки могли бы участвовать в цвето-пространственных взаимодействиях у человека, и несколько лет спустя совместно с А. Пирлменом он старательно, хотя и безуспешно, искал такие клетки в наружном коленчатом теле макака.
В конце 60-х годов и позже двойные оппонентные клетки изредка находили в коре обезьян, но их не связывали четко с какими-либо анатомическими структурами. Нам до сих пор непонятны некоторые из их особенностей. Например, у только что описанных клеток типа r+g– красное пятно, окруженное зеленым, часто вызывает слабую реакцию вместо энергичной, которой можно было бы ожидать.
Вперемешку с двойными оппонентными клетками обоих классов (красно-зелеными и желто-синими) встречались и обычные широкополосные клетки с центром и периферией. Эти широкополосные клетки отличались от клеток верхних слоев коленчатого тела и от клеток слоя 4C? коры более крупными размерами своих центров.
Мы с Маргарет Ливингстон предположили, что пузырьки представляют собой ветвь зрительного пути, имеющую дело с «цветом» в широком смысле, включая оттенки черного, белого и серого. Эта система, видимо, обособляется от остальной части зрительного пути либо в наружном коленчатом теле, либо в слое 4 стриарной коры. Коленчатое тело, возможно, имеет прямую, хотя и слабую проекцию на пузырьки. Кажется вероятной и проекция на них слоя 4C? — может быть, для них это даже главный источник входных сигналов. Проецируется ли на них слой 4C?, неясно. Обычно для реакции пузырьковых клеток необходим граничный контраст — нужны либо границы яркости, на которые отвечают широкополосные клетки с центром и периферией, либо цветоконтрастные границы, на которые реагируют двойные оппонентные клетки. Как уже говорилось, это равнозначно тому, что такие клетки играют роль в создании константности цвета.
Если пузырьковые клетки участвуют в константности цвета, они не могут осуществлять в точности те вычисления, которые рассматривались Лэндом, а именно сравнения между каким-то участком и его окружением для каждого из колбочковых спектральных поддиапазонов. Вместо этого они, по-видимому, осуществляют сравнение по типу Геринга: сопоставляют красно-зеленое в одном участке с красно-зеленым в окружающем фоне и делают то же самое для желто-синего и для интенсивности света. Но оба способа трактовки цвета — r, g и b в одном случае и b-w, r-g, и y-b в другом — по существу, эквивалентны. Цвет требует задания трех переменных: любому цвету соответствует тройка чисел, и мы можем представлять любой цвет точкой в трехмерном пространстве. Точки в таком пространстве можно представлять не единственным способом. Система координат может быть декартовой с осями, расположенными по нашему усмотрению, либо мы можем использовать полярные или цилиндрические координаты. В теории Геринга (в сетчатке и мозгу, наверное, тоже) просто используется иная совокупность осей для описания того же пространства. Эта теория, несомненно, упрощает действительность, так как пузырьковые клетки, принадлежащие к трем классам, вовсе не так сходны между собой, как горошины в стручке, — они значительно варьируют по относительной силе периферии и центра, по совершенству баланса между оппонентными цветами и по другим характеристикам, отчасти еще не вполне понятым. Сейчас мы можем лишь сказать, что физиология удивительно хорошо согласуется с психофизикой.
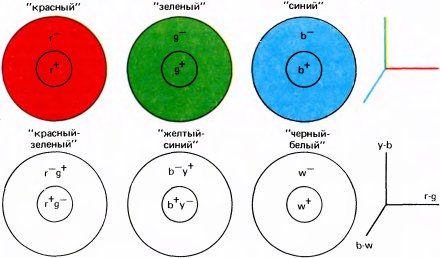
Рис. 130. Вверху: согласно формулировке Лэнда, константность цвета обусловлена наличием трех типов клеток, сравнивающих активацию данного набора колбочек (красных, зеленых или синих) в какой-то области сетчатки со средней активацией такого же набора в окружающей зоне. Результат — три числа, определяющие цвет данной области. Например, желтому, коричневому, темно-серому и оливковому соответствуют определенные тройки чисел. Мы можем, таким образом, отображать цвета в некотором цветовом пространстве с тремя осями, которые соответствуют красному, зеленому и синему.
Внизу: математически эквивалентная система, в которой тоже могут быть заданы три числа. Она, вероятно, ближе к тому способу, которым мозг определяет цвета. В любой точке сетчатки мы можем говорить о степени «красно-зелености», измеряемой неким прибором, регистрирующим относительную силу стимуляции красных и зеленых колбочек (и показывающим нуль для желтого и белого). Эта величина определяется для данной области, а ее среднее значение — для окружения; затем вычисляется отношение этих величин. Тот же процесс повторяется для желто-синей и черно-белой систем. Получаемые три числа достаточны для задания любого цвета.
Может возникнуть вопрос: зачем мозгу понадобились для отображения цвета такие на первый взгляд странные оси вместо более простой системы осей r, g и b, используемой в рецепторном слое сетчатки? По-видимому, цветовое зрение добавилось в ходе эволюции к «бесцветному» восприятию низших млекопитающих. Цветовое пространство у этих животных было одномерным, а колбочки разных типов (если животное обладало не одним типом колбочек) были объединены в общий пул. В ходе эволюции цветового зрения к уже существующей оси добавились еще две. Это было разумнее, чем отбросить уже сложившуюся черно-белую систему, а потом создавать три новые системы. Когда мы адаптируемся к темноте и используем только палочки, наше зрение лишается цвета и вновь отображается на единственную ось, в создании которой, очевидно, участвуют палочки. Это было бы не так просто сделать с осями r, g и b.
В настоящее время мы можем лишь гадать, какова схема связей двойных оппонентных клеток. На рис. 131 показан один из возможных вариантов их соединения с клетками верхних слоев коленчатого тела или с клетками слоя 4C? коры.
Излишне говорить, что все это чисто умозрительно; существенно лишь то, что двойная оппонентность каким-то образом должна достигаться: либо ее источник находится на нижних уровнях (сетчатки или коленчатое тело), и в этом случае мы просто еще не обнаружили клеток этого типа, либо это должно происходить в коре. Наличие таких клеток в сетчатке рыб не доказывает их существования у млекопитающих. Схема на рис. 131 всего лишь показывает, как могли бы быть сформированы центры рецептивных полей: для периферии мы всегда можем представить себе обратную процедуру с использованием клеток r–g+ коленчатого тела, поля которых широко перекрываются.
Наше стремление рассматривать цвет и форму как раздельные аспекты восприятия находит, таким образом, подтверждение в виде физического обособления пузырьковых и непузырьковых участков первичной зрительной коры. В областях выше стриарной коры — в зрительной зоне 2 и даже еще выше — эта обособленность сохраняется. Мы не знаем, где происходит их объединение, если оно вообще где-нибудь происходит.
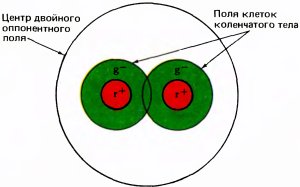
Рис. 131. Двойная оппонентная клетка может быть построена с помощью многих клеток мелкоклеточного слоя наружного коленчатого тела. Большая окружность на рисунке очерчивает центр рецептивного поля двойной оппонентной клетки; если клетка принадлежит к типу r+g–, то ее входами могут быть многочисленные клетки с центром r+ и периферией g– и с меньшей величиной центральной части поля. Аналогично, периферия у двойной оппонентной клетки может складываться из элементов типа r–g+.
Заключение
Самое интересное в цветовом зрении то, что здесь удается, объединив психофизические и нейрофизиологические методы, понять такие явления, как результаты смешения цветов или константность цвета, которые могли бы представляться совершенно загадочными. Проблемы, связанные с цветом, при всей их сложности, вероятно, проще тех, которые связаны с формой. Несмотря на открытие ориентационно-специфических клеток или клеток, реагирующих на концы линий, мы еще далеки от понимания нашей способности распознавать формы, отличать формы от их фона или воссоздавать трехмерную картину по плоским изображениям на сетчатке каждого глаза. Сопоставление этих двух модальностей — цвета и формы — вообще может ввести в заблуждение: вспомним, что одни лишь различия цвета на границах в отсутствие каких-либо различий освещенности могут приводить к восприятию формы. Цвет в этом смысле ничем не отличается от черного и белого и служит лишь одним из механизмов проявления формы.
Глаз, мозг, зрение
Дэвид Хантер Хьюбел; David Hunter Hubel;
Дек 30 2017