
Статьи. Предметно
Депривация и развитие
Депривация и развитие
До сих пор мы представляли себе мозг как некий полностью сформированный, зрелый механизм. Нас интересовало то, каким образом соединены между собой его части, как эти части функционируют в каждодневных ситуациях и как они обслуживают интересы животного. Все это, однако, оставляло открытым совершенно иной и чрезвычайно важный вопрос: как этот механизм создается?
В этой проблеме есть две главные составляющие. Развитие мозга в значительной части происходит до рождения животного, в материнской утробе. На первый взгляд мозг новорожденного младенца, хотя у него меньше складок и он несколько меньше мозга взрослого человека, в остальном не слишком от него отличается. Первый поверхностный взгляд, однако, вряд ли позволит выявить все подробности; ведь ребенок, понятно, не рождается со знанием алфавита или со способностью играть в теннис (или на арфе). Все эти достижения предполагают обучение, а под обучением мы, разумеется, понимаем формирование или модификацию нейронных связей под влиянием внешней среды. Таким образом, конечное состояние мозга — это результат как пренатального, так и постнатального развития. Во-первых, это развитие включает созревание как таковое, определяемое внутренними свойствами организма и происходящее до и после момента рождения; во-вторых, оно предполагает постнатальное созревание, определяемое тренировкой, обучением, образованием и опытом — все эти слова более или менее равнозначны.
Пренатальное развитие — тема гигантская; мне известно о нем не слишком много, и поэтому я не буду описывать его сколько-нибудь подробно. Один из самых интересных, но и самых трудных вопросов состоит в том, каким образом отдельные нервные волокна огромного пучка отыскивают место своего назначения. Глаз, коленчатое тело и кора, например, формируются независимо друг от друга; по мере их созревания растущие из них аксоны должны делать выбор из множества альтернатив. Волокно зрительного нерва должно прорасти через сетчатку к слепому пятну, затем пройти в составе зрительного нерва к хиазме и принять здесь решение о том, следует ли переходить на противоположную сторону; затем оно должно проследовать к наружному коленчатому телу выбранной стороны, подойти к нужному слою (или к области, которая позднее превратится в нужный слой), а затем в точности к нужной части этого слоя так, что полученная в результате топография станет надлежащим образом упорядоченной; и наконец, оно должно разветвиться, причем веточки должны подойти к надлежащим частям клетки коленчатого тела — к ее телу или к дендриту. Сходные требования предъявляются к волокну, растущему от наружного коленчатого тела к зоне 17 или от зоны 17 к зоне 18. Хотя этот общий аспект развития нейронов в настоящее время интенсивно изучается во многих лабораториях, мы еще не знаем, каким образом волокна отыскивают места своего назначения. Трудно даже угадать победителя среди нескольких основных конкурирующих факторов, к которым относятся механическая наводка, химические градиенты и взаимодействие с комплементарными молекулами вроде происходящего в иммунной системе. Многочисленные современные исследования как будто бы указывают на роль многих различных механизмов.
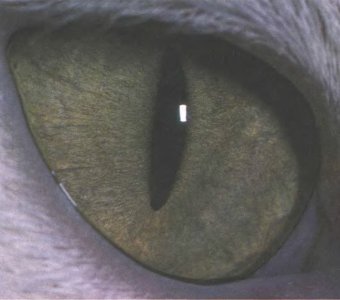
Рис. 132. По-видимому, щелевая форма зрачка, как у этой кошки и у многих ночных животных, позволяет более эффективно, чем круглый зрачок, снижать интенсивность света.
В этой главе мы рассмотрим в основном постнатальное развитие зрительной системы млекопитающих, особенно влияние, которое система испытывает со стороны окружающей среды. В отношении первых уровней зрительной системы кошки и обезьяны — сетчатки, коленчатого тела и, быть может, стриарной (первичной зрительной) коры — возникает естественный вопрос, сохраняется ли здесь после рождения некоторая пластичность. Я начну с описания простого эксперимента. Примерно к 1962 году стал известен ряд важных фактов о зрительной коре взрослой кошки: была открыта ориентационная избирательность, обнаружены «простые» и «сложные» клетки, о многих корковых нейронах стало известно, что они активируются сигналами от обоих глаз или проявляют различную степень глазодоминантности. Мы знали достаточно много о взрослом животном, чтобы задавать прямые вопросы относительно того, насколько пластична зрительная система. И вот Торстен Визел и я взяли котенка недельного возраста, когда глаза уже вот-вот должны открыться, и сшили его веки на одном глазу. Эта процедура может показаться жестокой, но она проводилась под наркозом, и проснувшийся котенок, возвращенный к своей матери и другим котятам, не выказал никаких признаков дискомфорта или страдания. Спустя десять недель мы хирургическим путем, опять-таки под наркозом, открыли глаз и стали регистрировать реакции корковых нейронов с целью выяснить, оказало ли отключение глаза какое-либо влияние на сам глаз или на зрительную систему.
Прежде чем я опишу результаты, я должен отметить, что идея этого эксперимента была подсказана длительной историей психологических исследований и наблюдений, связанных с клинической неврологией. Психологи широко экспериментировали со зрительной депривацией на животных в 40-х и 50-х годах, используя для оценки результатов поведенческие методы. Типичный эксперимент состоял в том, что животное с момента рождения содержали в полной темноте. Когда животных выносили на свет, они оказывались слепыми или по меньшей мере обладали неполноценным зрением. Слепота была в некоторой степени обратимой, но зрительные функции восстанавливались лишь постепенно и обычно не полностью.
Параллельно с этими экспериментами проводились клинические наблюдения над детьми с врожденной катарактой. Катаракта — это состояние, при котором хрусталик глаза мутнеет, он пропускает свет, но не позволяет формировать изображение на сетчатке. Катаракту у новорожденных, как и у взрослых, лечат путем хирургического удаления хрусталика и имплантации искусственной линзы или назначения очков с сильно выпуклыми стеклами. Это позволяет восстановить четкое изображение на сетчатке. Несмотря на относительную простоту операции, офтальмологи неохотно делали ее на очень маленьких детях или новорожденных, главным образом потому, что всякая операция, перенесенная в раннем детстве, статистически связана с большим риском, хотя этот риск и невелик. После удаления катаракты, скажем, в возрасте восьми лет и подбора очков результаты не оправдывали ожиданий. Зрение совсем не восстанавливалось: ребенок, как и раньше, оставался слепым, и глубокие дефекты сохранялись, несмотря на месяцы или годы попыток «научить его видеть». Ребенок, например, по-прежнему не мог отличить круг от треугольника. В результате появления и крушения надежд состояние ребенка вместо улучшения обычно ухудшалось. С этим резко контрастируют клинические случаи со взрослыми: у семидесятипятилетнего мужчины развивается катаракта на обоих глазах и он постепенно теряет зрение, но после трех лет слепоты катаракту удаляют, подбирают очки и зрение полностью восстанавливается. Зрение может даже улучшиться по сравнению с тем, что было до развития катаракты, поскольку хрусталик с возрастом желтеет и его удаление приводит к тому, что человек снова видит яркую синеву неба, которую воспринимают лишь дети и молодые люди.
По-видимому, зрительная депривация (лишение зрительного опыта) у детей приводит к пагубным последствиям, никогда не наблюдаемым у взрослых. Обычно психологи вполне резонно объясняли результаты своих экспериментов, а также клинические результаты неспособностью ребенка «научиться видеть» или (что, по-видимому, то же самое) невозможностью образования связей из-за нехватки тренирующего опыта.
Амблиопия — это частичная или полная потеря зрения, не связанная с какими-либо аномалиями самого глаза. Когда мы зашивали глаз у кошки или обезьяны, нашей целью было вызвать амблиопию, а затем выяснить, где в зрительном тракте возникнет аномалия. Результаты эксперимента с котенком поразили нас. Часто эксперимент дает неубедительные результаты, которые хороши лишь настолько, чтобы не опорочить идею полностью, но и не настолько определенны, чтобы вывести из них нечто полезное. Однако наш эксперимент явился исключением: результаты оказались совершенно ясными — когда мы открыли веки у котенка, сам глаз выглядел вполне нормальным; даже зрачок при его освещении сокращался обычным образом. Однако результаты регистрации активности корковых клеток никак нельзя было счесть нормальными. Хотя мы обнаружили много клеток с совершенно нормальными реакциями на ориентацию линий и их движение, мы нашли также, что вместо того, чтобы примерно половина клеток предпочитала один глаз другому, ни одна из двадцати пяти исследованных клеток не испытывала никакого влияния со стороны закрытого ранее глаза. (Пять клеток не испытывали влияния от обоих глаз, что изредка наблюдается даже у нормальных кошек.) Сравните это с нормальной кошкой, у которой около 15 % клеток монокулярны, причем около 7 % реагируют на левый глаз и 7 % — на правый. Гистограммы глазодоминантности для кошки, показанные вверху на рис. 133, позволяют сразу видеть это различие. Несомненно, что-то здесь основательно испортилось.
Вскоре мы повторили этот эксперимент на других котятах и на детенышах обезьян. На котятах более обширная серия опытов вскоре показала, что при сшивании век сразу после рождения в среднем лишь 15 процентов клеток предпочитают зашитый ранее глаз против примерно 50 процентов у интактных котят. На обезьянах были получены примерно такие же результаты (см. нижние гистограммы на рис. 133). Среди немногочисленных клеток, отвечавших на стимуляцию закрытого ранее глаза, многие казались аномальными; они реагировали вяло, быстро утомлялись, и у них отсутствовала точная ориентационная настройка.
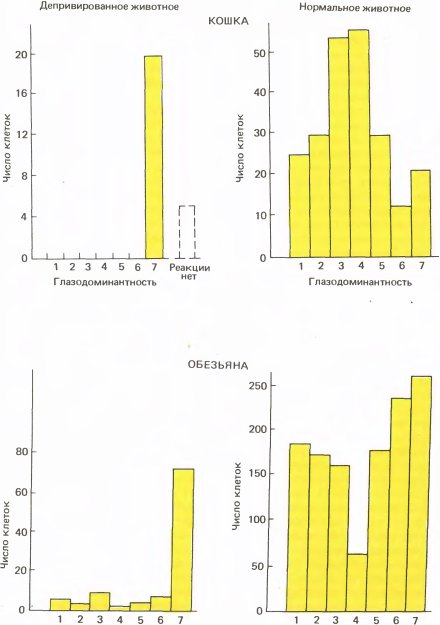
Рис. 133. Правый глаз котенка был на два с половиной месяца лишен зрительных стимулов в возрасте примерно десяти дней, когда глаза обычно уже открыты. В этом эксперименте (верхние гистограммы) мы зарегистрировали ответы только двадцати пяти клеток. (В последующих экспериментах мы смогли исследовать больше клеток и обнаружили, что небольшая доля их испытывала влияние со стороны ранее закрытого глаза.) В опытах на детеныше обезьяны (нижние гистограммы) результаты оказались весьма сходными. Его правый глаз закрыли в возрасте двух недель и держали закрытым на протяжении 18 месяцев. Позднее мы нашли, что те же результаты получаются и при закрытии глаза всего на несколько недель.
Результат, подобный этому, ставит множество вопросов. Где в зрительной системе возникала аномалия? В глазу? В коре? Могла ли кошка видеть закрытым ранее глазом, несмотря на корковую аномалию? Вызывалась ли эта аномалия просто световой депривацией или же лишением возможности видеть различные формы? Имел ли значение возраст, в котором мы закрывали глаз? Была ли аномалия результатом неиспользования глаза или она обусловливалась какими-то иными причинами? Чтобы все это выяснить, потребовалось много времени, но мы можем сформулировать результаты в нескольких словах.
Для того чтобы определить локализацию дефекта, нужно было, очевидно, регистрировать реакции от более низких уровней, начиная, скажем, с сетчатки или с коленчатого тела. Результаты оказались однозначными: как в сетчатке, так и в коленчатом теле было множество клеток с практически нормальными реакциями. Клетки тех слоев коленчатого тела, к которым шли волокна от ранее закрытого глаза, имели обычные рецептивные поля с центром и периферией; они хорошо реагировали на маленькие пятна и плохо — на рассеянный свет. Единственным намеком на аномалию была некоторая вялость реакций этих клеток по сравнению с клетками слоев, получавших входные сигналы от нормального глаза.
Зная об этой относительной нормальности, мы были поражены, впервые увидев под микроскопом наружное коленчатое тело, окрашенное по Нисслю. Его аномальность была видна практически и без микроскопа. Коленчатое тело у кошки организовано несколько проще, чем у обезьяны; оно состоит в основном из двух крупноклеточных слоев, расположенных наверху, а не внизу, как у обезьяны. Верхний слой имеет вход от контралатерального, а нижний — от ипсилатерального глаза. Под этими слоями лежит довольно плохо выраженный слой мелких клеток с несколькими подразделениями, которые я здесь игнорирую. На каждой стороне крупноклеточный слой, получавший входы от закрытого глаза, был бледнее и явно тоньше своего партнера, который выглядел совершенно нормальным. Клетки аномальных слоев были не только бледными, но и сморщенными примерно до двух третей своего нормального поперечного сечения. Результат, полученный при закрытии правого глаза, показан на рис. 134. Сходные результаты были получены и на макаках (рис. 135). Таким образом, мы столкнулись с парадоксом, на разрешение которого потребовалось несколько лет: в клетках наружного коленчатого тела, физиологически казавшихся сравнительно нормальными, гистологическое исследование показывало явную патологию. Как бы то ни было, мы теперь имели ответ на наш исходный вопрос, поскольку корковые клетки, практически не отвечавшие на стимуляцию ранее закрытого глаза, тем не менее, видимо, получали значительный и как будто бы нормальный поток входных сигналов от коленчатого тела; это позволяло думать, что первичное повреждение локализуется не в глазу или коленчатом теле, а главным образом в коре. При гистологическом исследовании коры мы не находили абсолютно никаких признаков аномалии. Как мы увидим далее, в коре все же были анатомические дефекты, но они не выявлялись с помощью применявшихся нами методов окраски.
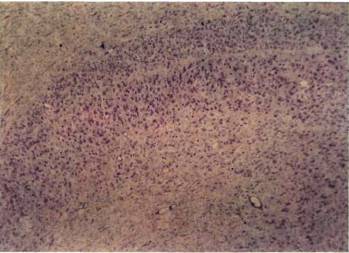
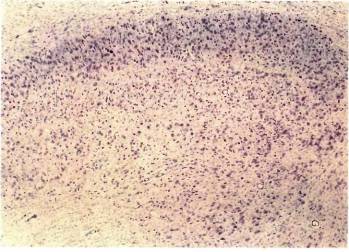
Рис. 134. Если правый глаз котенка закрыть в возрасте 10 дней на три с половиной месяца, то в его наружных коленчатых телах возникнут явные аномалии. В верхних частях обеих микрофотографий можно видеть два главных слоя. Вверху: верхний слой на левой стороне (контралатеральной по отношению к закрытому правому глазу) утончен и бледно окрашен. Внизу: на правой (ипсилатеральной) стороне аномален нижний из двух слоев. Оба слоя вместе имеют толщину около 1 мм. Гистограмма глазодоминантности показана на рис. 133.
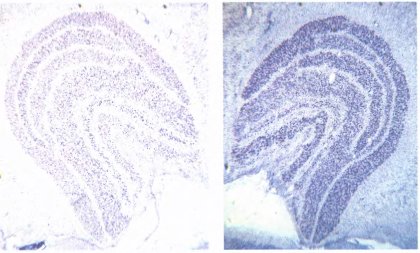
Рис. 135. Поперечные срезы левого и правого наружных коленчатых тел обезьяны, правый глаз которой был закрыт в возрасте двух недель на 18 месяцев. Видны аномальные слои, они выглядят более бледными. На обеих сторонах это те слои, которые получают входные волокна ранее закрытого (правого) глаза,— слои 1, 4 и 6 слева, 2, 3 и 5 справа (счет идет снизу вверх).
Клетки измененных слоев мельче, но это невозможно рассмотреть при таком малом увеличении. Ширина всей структуры около 5 миллиметров.
Следующий вопрос заключался в том, что же именно порождает аномалию при закрытии глаза. Сшивание век уменьшает количество света, достигающего сетчатки, приблизительно в 10–50 раз; оно, разумеется, препятствует также созданию на сетчатке каких-либо изображений. Может ли аномалия вызываться просто уменьшением количества света? Чтобы выяснить это, мы вставляли в один глаз новорожденного котенка матовую контактную линзу из пластмассы, по консистенции близкой к материалу мячика для настольного тенниса. У других животных мы вместо этого пришивали на одном глазу тонкую просвечивающую мембрану, играющую роль дополнительного века и называемую мигательной перепонкой, которая имеется у кошек, но отсутствует у людей. Пластмасса или перепонка уменьшала количество света всего лишь вдвое, но предотвращала формирование каких-либо сфокусированных изображений. Результаты оказались такими же: аномальная корковая физиология и аномальная гистология коленчатого тела. Очевидно, повреждение вызывалось невозможностью воспринимать формы, а не просто недостатком света.
У нескольких котят перед регистрацией нейронных ответов мы тестировали зрение, надевая непрозрачную черную контактную линзу на тот глаз, который не был закрыт, и затем наблюдая за поведением животного. Животные были, несомненно, слепы на депривированный глаз: будучи помещены на низком столике, они уверенно шли к его краю, перешагивали его и падали на разложенную на полу подстилку. Ходя по полу, они обычно натыкались на ножки стола. Этого никогда не делает ни одна нормальная, уважающая себя кошка. Аналогичные тесты с незакрытым глазом показали, что зрение было совершенно нормальным.
Далее мы провели длительные исследования на кошках и обезьянах с целью выяснить, зависят ли результаты от возраста, в котором закрывают глаз, и от продолжительности депривации. Вскоре выяснилось, что возраст имеет решающее значение. У взрослой кошки, у которой один глаз был отключен более чем на год, не наблюдалось ни слепоты на этот глаз, ни потери корковых реакций, ни патологии коленчатого тела. (Первая депривированная кошка, мать нашего первого помета котят, была взрослой по определению!) После многочисленных экспериментов мы пришли к выводу, что где-то между рождением и зрелым возрастом должен существовать период пластичности, когда депривация приводит к корковому дефекту. Как выяснилось, у кошки этот критический период длится от четвертой недели до четвертого месяца. Неудивительно, что закрытие глаза мало влияет до наступления четвертой недели, так как в первый месяц жизни кошка почти не пользуется зрением: глаза открываются лишь около 10-го дня, а несколько последующих недель котята со своей матерью прячутся за диваном. Чувствительность к депривации быстро возрастает и достигает максимума в первые недели критического периода. В это время отключение одного глаза даже на несколько дней приводит к заметному искажению гистограммы глазодоминантности. В последующие четыре месяца время депривации, необходимое для получения явных эффектов, неуклонно возрастает; иными словами, чувствительность к депривации уменьшается и сходит на нет.
На рис. 136 представлены гистограммы, иллюстрирующие некоторые результаты, полученные на обезьянах. На левом графике — эффект отключения в возрасте пяти дней одного глаза на 6 недель; почти нет клеток, которые отвечали бы на стимуляцию ранее закрытого глаза. Гораздо более кратковременная депривация (средний график) тоже вызывает сильно выраженный эффект, но явно меньший, чем при более длительной депривации. В возрасте четырех месяцев чувствительность снижается настолько, что даже отключение глаза на 5 лет (правый график) дает эффект хотя и весьма заметный, но все же несравнимый с последствиями более ранней депривации.
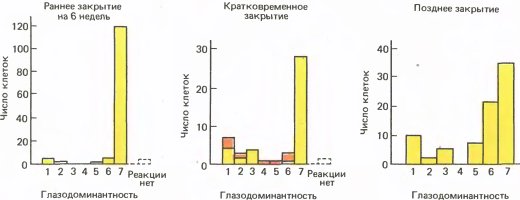
Рис. 136. Слева: у обезьяны, правый глаз которой был зашит в возрасте пяти дней на шесть недель, практически полностью доминирует левый глаз. В середине: закрыть глаз всего на несколько дней в возрасте нескольких недель уже достаточно, чтобы вызвать заметный сдвиг глазодоминантности. Более темным цветом показано число аномальных клеток. Справа: если закрытие глаза отложить до четырехмесячного возраста, то очень длительное его отключение (в данном случае на пять лет) приводит к гораздо меньшему сдвигу глазодоминантности, чем кратковременное отключение в возрасте нескольких недель.
При изучении периода чувствительности у кошек и обезьян были получены весьма сходные результаты. У обезьян этот период начинается раньше, в момент рождения, а не в возрасте четырех недель, и длится дольше, постепенно заканчиваясь к году, а не к четвертому месяцу. Чувствительность наиболее высока в первые две недели жизни, на протяжении которых всего нескольких дней депривации оказывается достаточным, чтобы вызвать выраженный сдвиг глазодоминантности. Отключение глаза у взрослой обезьяны независимо от его продолжительности не вызывает никаких вредных последствий. У одной взрослой обезьяны мы закрывали глаз на пять лет, после чего не отмечалось ни слепоты, ни коркового дефекта, ни сморщивания клеток в коленчатых телах.
Восстановление
Следующим интересовавшим нас вопросом был вопрос о возможности какого-либо восстановления нарушенной функции у обезьяны после открытия отключенного ранее глаза. Оказалось, что никакого или почти никакого физиологического восстановления не происходит, если глаз, закрытый на неделю или на больший срок, просто открыть и ничего более не делать. Даже спустя несколько лет кора оставалась приблизительно такой же аномальной, как и в момент открытия глаза (рис. 137, левый график). Если же, открыв один глаз, закрывали другой, ранее открытый (эту процедуру мы назвали реверсией), то восстановление наблюдалось, но лишь в том случае, если обезьяна все еще находилась в критическом периоде (рис. 137, средний и правый графики — для случая ранней и поздней реверсии). После окончания критического периода даже реверсия — закрытие другого глаза на несколько лет — не давала практически ничего, кроме незначительного восстановления структуры или функции.
Способность обезьяны видеть не всегда соответствовала физиологическому состоянию зрительной коры. Без реверсии зрение в ранее закрытом глазу никогда не восстанавливалось. При реверсии зрение возвращалось и часто достигало почти нормального уровня, причем это происходило даже в случаях поздней реверсии, когда физиология первоначально закрытого глаза оставалась весьма аномальной. Мы все еще не можем объяснить это несоответствие между отсутствием существенной физиологической или анатомической нормализации и тем, что в ряде случаев зрение, по-видимому, в значительной степени восстанавливалось. Возможно, что те и другие наблюдения отражают разные стороны зрительной функции. Мы испытывали остроту зрения, измеряя такие показатели, как наименьший различимый разрыв в линии или окружности. Такое тестирование, однако, может не давать полного представления о зрительной функции. Трудно поверить, что столь выраженные физиологические и анатомические дефекты проявляются на поведенческом уровне всего лишь в незначительном снижении остроты зрения.
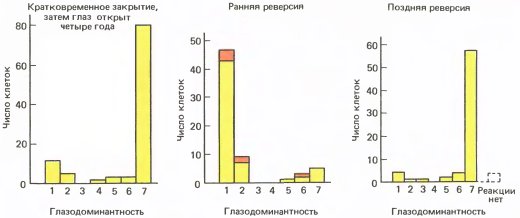
Рис. 137. Слева: у макака один глаз закрыли при рождении на 9 дней, а затем открыли. Ответы клеток регистрировали по прошествии четырех лет, на протяжении которых зрение животного многократно тестировалось. Даже столь длительный период, когда оба глаза были открыты, не привел к заметному восстановлению физиологических показателей.
В середине: правый глаз был отключен с рождения; через пять с половиной недель этот глаз открыли, а левый закрыли. Когда в возрасте шести месяцев регистрировали ответы в правом полушарии, в отношении большинства клеток резко доминировал первоначально закрытый правый глаз.
Справа: в этом случае правый глаз закрыли в возрасте 7 дней на один год, после чего открыли его и закрыли левый глаз. Спустя еще год открыли и левый глаз, после чего оба глаза оставались открытыми. Когда, наконец, в возрасте трех с половиной лет регистрировали корковые ответы, большинство клеток реагировало на сигналы от того глаза, который был открыт в самом начале. Очевидно, один год — слишком поздний срок для реверсии.
Природа повреждений
Описанные выше результаты ясно показали, что отсутствие изображений на сетчатке в раннем периоде жизни ведет к глубоким и стойким нарушениям корковой функции. Оставались, однако, открытыми два важных вопроса относительно природы этого явления. Первым из них был вопрос, связанный с дилеммой «природа или воспитание»: лишали ли мы животных индивидуального опыта, в котором они нуждались для создания правильных связей, или же мы разрушали те связи, которые уже существовали, были заранее сформированы и функционировали при рождении животного? Практически все эксперименты с выращиванием животных в темноте, проведенные за десятки лет до нашей работы, интерпретировались в контексте проблемы обучаемости. Кору головного мозга, которую большинство людей считали (и продолжают считать) физиологическим субстратом памяти и мышления, рассматривали как нечто подобное запоминающему устройству емкостью в 1 мегабайт, за которое мы так дорого платим при покупке компьютера: и в том и в другом содержится множество элементов и соединений, но нет никакой информации, пока мы ее туда не заложим. Короче говоря, кору считали своего рода tabula rasa.

Рис. 138. Японский макак (Macaca fuscata), самый крупный из всех макаков, обитает на земле и на деревьях в северной части Японии. Он защищен от холода густым серовато-бурым мехом.
Одним из очевидных подходов к решению поставленного вопроса могла бы быть прямая регистрация активности нейронов у новорожденных котят или обезьян. Если бы для создания нужных связей требовалось обучение, мы не обнаружили бы ничего похожего на ту высокоразвитую специфичность, которая наблюдается у взрослых животных. Отсутствие специфичности, однако, не решило бы проблему, так как неразвитость связей можно было бы объяснить либо незрелостью (тем, что построение генетически запрограммированной системы связей еще не завершилось), либо отсутствием сенсорного опыта. С другой стороны, обнаружение такой специфичности говорило бы против механизма обучения. Мы не ожидали, что эксперименты на котятах будут простыми; так оно и оказалось. Котенок рождается с очень незрелой зрительной системой и совсем не использует глаза примерно до десятого дня, когда они открываются. В это время даже среды глаза, прозрачные субстанции, находящиеся между роговицей и сетчаткой, еще не вполне прозрачны и не позволяют получить четкое изображение на сетчатке. Незрелая зрительная кора действительно реагировала вяло, несколько непредсказуемо и в целом сильно отличалась от нормальной зрительной коры взрослой кошки; тем не менее мы нашли много клеток с явной ориентационной специфичностью. Чем больше времени проходило от рождения до регистрации, тем большее число клеток по своему поведению приближалось к клеткам взрослого типа — может быть, из-за прояснения глазных сред и общего укрепления организма, а быть может, и в результате обучения. Различные группы исследователей объясняют это по-разному.
Наиболее убедительные данные были получены на новорожденных обезьянах. У макака на другой день после рождения зрительная система обнаруживает поразительную зрелость: детеныши смотрят в разные стороны, следят взором за предметами и проявляют большой интерес к окружающему (рис. 139). В соответствии с таким поведением клетки первичной зрительной коры у них проявляют столь же острую ориентационную настройку, как и у взрослой особи. Последовательно расположенные клетки хорошо упорядочены по ориентации (см. график на рис. 140). Хотя мы и отмечали некоторые различия между новорожденными и взрослыми животными, система ориентационно-специфичных рецептивных полей, эта отличительная особенность стриарной коры, казалась вполне четко организованной.

Рис. 139. На следующий день после рождения макак-детеныш осматривается, фиксирует взором предметы, проявляет живой интерес к окружающему миру. У человека и кошки зрительная функция достигает такой степени зрелости лишь спустя много недель после рождения.
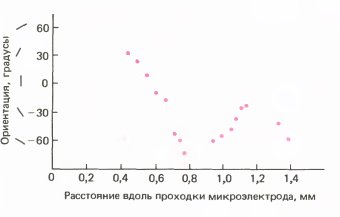
Рис. 140. У новорожденного макака клетки коры проявляют примерно такую же ориентационную избирательность, как и у взрослых особей, и последовательность ориентаций почти так же упорядоченна.
У макака (в сравнении с кошкой или человеком) зрительная система новорожденной особи может считаться весьма зрелой, но она, несомненно, анатомически отличается от зрительной системы взрослой обезьяны. Срезы коры, окрашенные по Нисслю, выглядят различно: у детенышей более тонкие слои и более плотная упаковка клеток. Ле-Вэй впервые показал, что даже общая площадь стриарной коры от рождения к зрелому возрасту увеличивается примерно на 30 процентов. Если окрасить кору по методу Гольджи или исследовать ее под электронным микроскопом, отличия станут еще более очевидными: у клеток обычно менее развита система дендритов и меньше синапсов. Ввиду этих различий было бы удивительно, если бы кора уже при рождении функционировала в точности так же, как у взрослого животного. С другой стороны, через месяц после рождения дендриты и синаптические связи все еще не достигают полного развития. Вопрос «природа или воспитание?» означает: определяется ли постнатальное развитие опытом или все еще протекает по врожденной программе? Мы пока не уверены в ответе, но, судя по сравнительной нормальности реакций корковых клеток при рождении, можно сделать вывод, что нарушение их после депривации обусловлено в основном «порчей» связей, имевшихся к моменту рождения, а не невозможностью их формирования без индивидуального опыта.
Второй важный вопрос касается причины этой порчи. На первый взгляд ответ казался почти очевидным. Мы полагали, что деградация связей обусловлена их неиспользованием, подобно тому как происходит атрофия мышц ноги при фиксации колена или лодыжки гипсовой повязкой. В сморщивании клеток коленчатого тела мы видели проявление постсинаптической атрофии — так называют уменьшение величины клеток в наружных коленчатых телах взрослых животных или людей после удаления глаза. Эти предположения оказались неверными. Мы считали их настолько самоочевидными, что я не уверен, что мы когда-либо подумали бы о проведении специального эксперимента для их проверки. Нам пришлось изменить свою точку зрения только в результате одного необязательного, как нам тогда казалось, эксперимента, причин постановки которого я уже не помню.
Мы зашили оба глаза сначала у новорожденного котенка, а потом у новорожденного детеныша обезьяны. Если бы нарушение реактивности корковых клеток, получающих входные сигналы от одного глаза, было обусловлено их бездействием, то отключение обоих глаз удвоило бы дефект: мы практически не смогли бы обнаружить клеток, реагирующих на левый или правый глаз. На самом же деле, к нашему великому изумлению, мы не получили ничего похожего на отсутствие клеточных реакций: после открытия глаз добрая половина клеток стриарной коры снова отвечала нормально, одна четверть отвечала аномально и одна четверть не реагировала вовсе. Мы вынуждены были заключить, что невозможно предсказать судьбу корковой клетки в ситуации, когда закрыт один глаз, если неизвестно, был ли при этом закрыт и другой. Отключите один глаз, и клетка почти наверняка утратит свои связи с ним; но отключите оба глаза, и у вас будут неплохие шансы на то, что связь сохранится. Мы, очевидно, столкнулись здесь не с бездействием клеток и их связей, а с какого-то рода конкуренцией между двумя глазами. Дело обстоит так, как если бы клетка вначале имела две группы синаптических входов — два входных пути, по одному от каждого глаза, но при неиспользовании одного пути одерживает верх другой, завладевая территорией первого (рис. 141).
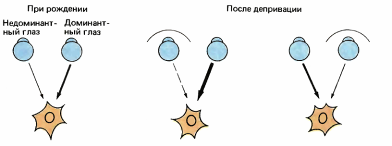
Рис. 141. Мы предполагаем, что корковая клетка имеет входы от двух источников, по одному от каждого глаза, и что закрытие одного глаза ослабляет связи от этого глаза и усиливает связи от другого.
Такое рассуждение, мы полагали, вряд ли применимо к сморщиванию клеток коленчатого тела, поскольку эти клетки монокулярны и здесь не видно каких-либо возможностей конкуренции. В то время мы не смогли объяснить атрофию клеток в слоях, соответствующих закрытому глазу. При закрытии обоих глаз сморщивание клеток коленчатого тела не столь заметно, но здесь трудно что-либо утверждать, так как при этом отсутствуют нормальные слои, которые можно было бы использовать для сравнения. Наше понимание всей этой проблемы не продвинулось, пока мы не начали использовать некоторые новые методы экспериментальной анатомии.
Косоглазие
Наиболее частой причиной амблиопии у людей бывает косоглазие, или страбизм — непараллельность оптических осей глаз. Косоглазие может быть сходящимся или расходящимся. Причина косоглазия неизвестна, почти наверняка существует несколько причин. Иногда косоглазие возникает после рождения, в первые месяцы жизни, когда глаза едва-едва начинают фиксировать и прослеживать предметы. Нарушения прямоты взгляда могут быть результатом аномалий глазных мышц или механизмов мозгового ствола, обслуживающих движение глаз.
У некоторых детей косоглазие, по-видимому, связано с дальнозоркостью. Для правильной фокусировки удаленного объекта хрусталик дальнозоркого глаза должен принять столь же выпуклую форму, как хрусталик нормального глаза, когда он фокусирует близкий объект. Округление хрусталика при рассматривании близких предметов означает сокращение цилиарной мышцы внутри глаза; этот процесс называют аккомодацией. Когда человек с нормальным зрением осуществляет аккомодацию на близкое расстояние, глаза одновременно автоматически поворачиваются к срединной плоскости (конвергенция). Эти два процесса показаны на рис. 142. Нейронные сети мозгового ствола, управляющие аккомодацией и конвергенцией, вероятно, связаны друг с другом и, возможно, перекрываются; как бы то ни было, трудно осуществить одну из этих реакций без другой. Когда дальнозоркий человек осуществляет аккомодацию, которая необходима ему для фокусировки даже удаленного объекта, один или оба глаза могут повернуться внутрь, хотя конвергенция в этом случае нежелательна. Если дальнозоркий ребенок не носит очки, поворот глаза может стать со временем постоянной привычкой. Это объяснение косоглазия, конечно, справедливо лишь в некоторых, но не во всех случаях, поскольку косоглазие не всегда сопровождается дальнозоркостью, а у некоторых людей с косоглазием один глаз поворачивается кнаружи, а не вовнутрь.
Косоглазие можно излечить хирургическим путем, изменив места прикрепления наружных мышц глаза. Операция обычно с успехом исправляет положение глазных осей, но до последнего десятилетия ее чаще всего откладывали до тех пор, пока ребенок не достигнет возраста четырех — десяти лет (по той же причине, по которой откладывали удаление катаракты, — чтобы несколько уменьшить риск операции).
Косоглазие, возникающее у взрослых, например вследствие повреждения нерва или глазной мышцы, сопровождается, конечно, двоением в глазах. Чтобы испытать это состояние, достаточно лишь слегка надавить на один глаз снизу и сбоку. Двоение в глазах может быть весьма неприятным и даже выводить человека из строя, и если никакое лучшее решение невозможно, один глаз следует закрыть повязкой. В противном случае двоение в глазах останется до тех пор, пока не будет исправлено косоглазие. У детей с косоглазием, однако, двоение в глазах редко продолжается длительное время; оно сменяется либо поочередным использованием двух глаз, либо подавлением зрения в одном глазу.
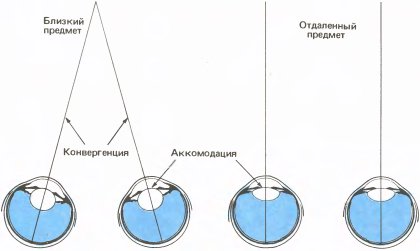
Рис. 142. При рассматривании близкого предмета происходят две приспособительные реакции: хрусталик округляется в результате сокращения цилиарных мышц, и глаза поворачиваются к срединной плоскости головы.
Когда ребенок чередует глаза, он фиксирует предметы сначала одним глазом, в то время как нефиксирующий глаз поворачивается внутрь или наружу, а затем другим глазом, а первый при этом отводится в сторону. (Чередующееся косоглазие весьма распространено, и если вы знаете его признаки, вы легко распознаете его.) Глаза используются по очереди, иногда смена происходит почти ежесекундно, и пока один глаз смотрит, другой как бы ничего не видит. В любой момент, когда один глаз расположен правильно, а другой отклоняется, говорят, что зрение в отклоненном глазу подавлено. Подавление знакомо всякому, кто привык смотреть в монокулярный микроскоп, прицеливаться из ружья или выполнять какую-либо иную задачу, используя только один глаз при открытом другом глазе. Для подавленного глаза зрительное восприятие попросту исчезает. Ребенок с чередующимся косоглазием всегда подавляет тот или другой глаз, но если проверить зрение в каждом глазу по отдельности, то обычно окажется, что оба глаза вполне нормальны.
Некоторые дети с косоглазием не чередуют глаза, а используют все время один глаз при подавленном другом. Если один глаз привычно подавляется, то зрение в подавляемом глазу проявляет тенденцию к ухудшению. Острота зрения падает, особенно в области центральной ямки и около нее, и, если это состояние длится достаточно долго, глаз может стать практически слепым. Такого рода слепоту офтальмологи называют amblyopia ex anopsia. Это наиболее распространенный вид амблиопии и даже слепоты вообще.
Естественно, что нам пришло в голову попытаться вызвать косоглазие и, следовательно, амблиопию у котят или обезьяны путем хирургической перерезки одной из мышц глаза после рождения, так как при этом мы смогли бы провести физиологические исследования и посмотреть, какая часть зрительного пути нарушена. Мы сделали это на полудюжине котят и к своему неудовольствию обнаружили, что у котят, как и у многих детей, развилось чередующееся косоглазие; они смотрели сначала одним глазом, а потом другим. Протестировав каждый глаз в отдельности, мы вскоре убедились, что в обоих глазах зрение было нормальным. Стало ясно, что нам не удалось вызвать амблиопию, и мы начали думать, что же делать дальше. Мы решили провести на одном из котят регистрацию клеточных реакций, хотя совершенно не представляли себе, что можно было таким образом выяснить (исследования часто включают поиски «ощупью»). Результаты оказались совершенно неожиданными. Изучая клетку за клеткой, мы вскоре поняли, что с мозгом случилось нечто странное: каждая клетка реагировала совершенно нормально, но только на стимуляцию одного глаза. По мере продвижения электрода в коре шел ряд нейронов, активируемых левым глазом, затем этот ряд обрывался и в действие вступал другой глаз. В отличие от того, что мы наблюдали после закрытия глаза, ни один глаз не был ущемлен в своих правах по сравнению с другим в общем балансе доминирования. Иногда в зоне перехода появлялись бинокулярные клетки, но, как видно из рис. 143, поля бинокулярных клеток во всей популяции составляли около 20 процентов вместо 85 в норме.
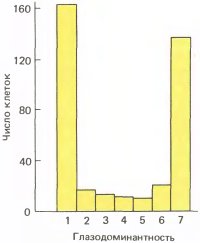
Рис. 143. Если у новорожденного котенка перерезать одну из наружных мышц глаза, спустя три месяца подавляющее большинство клеток оказываются монокулярными, попадая в группы 1 и 7.
Мы заинтересовались тем, что произошло с исходно бинокулярными клетками — может быть, они просто погибли или совсем утратили способность реагировать, так что оставались только первично монокулярные клетки? Это казалось весьма маловероятным, так как по мере продвижения электрода мы находили обычное изобилие реагирующих клеток — не было ничего похожего на пятикратное уменьшение их общего числа. У нормальной кошки при горизонтальном продвижении электрода в верхних слоях коры мы обычно встречаем примерно от 10 до 15 клеток подряд с доминированием одного и того же глаза; все они явно принадлежат к одной и той же колонке глазодоминантности, и две или три из них могут быть монокулярными. У косоглазых котят мы точно так же наблюдали по 10–15 клеток с доминированием одного глаза, но теперь все, кроме двух или трех, были монокулярными. Видимо, каждая клетка полностью или почти полностью перешла в ведение того глаза, который ранее просто оказывал на нее большее влияние.
Чтобы оценить значение этого результата, следует вспомнить, что мы в сущности никак не изменили общее количество зрительных стимулов, получаемых каждой сетчаткой. Поскольку у нас не было оснований полагать, что мы повредили какой-либо из двух глаз, мы пришли к выводу, что общий поток импульсов в двух зрительных нервах должен был остаться нормальным.
Каким же образом косоглазие смогло вызвать столь радикальные изменения корковой функции? Чтобы ответить на этот вопрос, нам нужно рассмотреть, как два глаза взаимодействуют друг с другом в норме. Косоглазие изменило связь между стимулами, воздействующими на оба глаза. Когда мы рассматриваем какой-то объект, изображения любой его точки в норме попадают на те места двух сетчаток, которые находятся на одинаковом расстоянии и в одинаковом направлении относительно обеих центральных ямок, — они попадают на корреспондирующие точки. Если изображение на левой сетчатке активирует какую-то бинокулярную клетку (это происходит, если рецептивное поле клетки пересекается границей темнота/свет, ориентация которой в точности соответствует данной клетке), то эта клетка будет также активироваться изображением на правой сетчатке. Это обусловлено тремя причинами:
1) изображения попадают на одни и те же участки обеих сетчаток;
2) бинокулярная клетка (если только она не специализирована для восприятия глубины) имеет рецептивные поля точно в одних и тех же участках обеих сетчаток;
3) ориентационные предпочтения бинокулярных клеток всегда близки для обоих глаз. Очевидно, что в случае косоглазия причина 1 отпадает: если при несовпадающих изображениях один глаз в данный момент побуждает клетку разряжаться, то будет ли делать то же самое другой глаз, зависит только от случая. Для одиночной клетки это, по-видимому, единственный фактор, изменяющийся при косоглазии. Когда котенок несколько недель или месяцев находится в такой ситуации, при которой сигналы от обоих глаз более не соответствуют друг другу, это приводит к тому, что более слабая из двух групп входных связей данной клетки становится еще слабее и часто практически исчезает. Таким образом, мы имеем здесь пример патологических изменений, вызванных не отсутствием стимуляции, а всего лишь разрывом нормальных временны?х соотношений между двумя совокупностями стимулов, — изощренный удар, если учитывать серьезность последствий.
В аналогичных экспериментах на обезьянах были получены такие же результаты; поэтому кажется, что косоглазие приводит к тем же последствиям и у людей. По клиническим данным, у человека с длительным чередующимся косоглазием даже после устранения этого дефекта обычно не восстанавливается способность к восприятию глубины. Хирург может привести оба глаза в соответствие лишь с точностью до нескольких градусов, а больной, вероятно, уже утратил способность достигать полного слияния двух изображений путем правильной установки глаз с точностью до нескольких угловых минут. Хирургическое исправление косоглазия выравнивает глаза достаточно хорошо, и нормальные нейронные механизмы справились бы с оставшимися несколькими градусами тонкой подстройки, но при косоглазии именно эти механизмы, включающие бинокулярные клетки коры, оказываются нарушенными. Для восстановления, вероятно, потребовался бы длительный период точного выравнивания глаз, который предполагает как нормальную мышечную подстройку, так и подстройку, определяемую бинокулярным зрением.
Все эти соображения, объясняющие сдвиг глазодоминантности, весьма напоминают модель, позволяющую объяснить ассоциативное научение изменениями в синапсах. Эта модель была предложена психологом Дональдом Хеббом из Университета Мак-Гилла. Ее основная идея состоит в том, что синапс между двумя нейронами A и C становится тем более эффективным, чем чаще за возбуждением нейрона A следует возбуждение нейрона C, независимо от того, чем это последнее будет вызвано (рис. 144). Таким образом, для повышения эффективности синапса вовсе не обязательно, чтобы разряд нейрона C был следствием разряда A. Предположим, например, что с C образует синапс еще один нейрон, B, причем синапс A–>C слаб, а синапс В–>C весьма эффективен; допустим далее, что A и B разряжаются почти одновременно или же B чуть раньше A и что C дает разряд не под влиянием A, а в результате сильного воздействия B. В модели Хебба уже сам факт, что C разряжается сразу после A, повышает эффективность синапса А–>С. Мы предположим также, что если импульсы, приходящие к C от A, не сопровождаются импульсами в C, то синапс A–>C ослабевает.
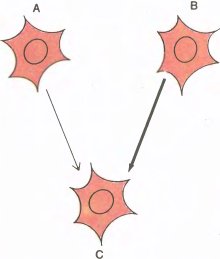
Рис. 144. Клетка C получает входы от клетки левого глаза A и от клетки правого глаза B. Согласно модели синапса Хебба, если разряды клетки C следуют за разрядами клетки A, то такая последовательность событий стремится усилить синапс A–>C.
Применим эту модель к бинокулярной конвергенции у нормального животного. Пусть клетка C — бинокулярная, аксон A идет от не доминантного, а аксон B — от доминантного глаза. Недоминантный глаз будет реже активировать клетку C, чем доминантный. Согласно гипотезе Хебба, синапс A–>C будет сохраняться или усиливаться, пока за импульсами в A будут следовать импульсы в C, а такая последовательность более вероятна, если в надлежащий момент времени от другого глаза по аксону B постоянно поступает «подкрепление». А именно это и будет происходить при точном совмещении изображений на обеих сетчатках. Если активность в A не сопровождается активностью в C, синапс A–>C за длительный период времени ослабеет. Получить прямое доказательство, что модель Хебба применима к косоглазию, — дело нелегкое (по крайней мере в ближайшем будущем), но сама идея, мне кажется, заслуживает внимания.
Анатомические последствия депривации
То, что в коленчатых телах, где нет или почти нет возможностей для конкуренции глаз, мы не смогли обнаружить заметных физиологических дефектов, казалось, подтверждает идею о том, что последствия закрытия одного глаза связаны с конкуренцией, а не с бездействием глаза. Правда, клетки коленчатого тела были атрофичными, но — рассуждали мы — вряд ли можно ожидать, что все будет ясно. Если конкуренция и в самом деле играет важную роль, то корковый слой 4C мог, как нам казалось, быть подходящим объектом для проверки этой идеи, поскольку клетки в нем тоже монокулярны и конкуренция маловероятна, так что чередование полосок, связанных с левым и с правым глазом, должно остаться ненарушенным. Итак, путем длинных микроэлектродных проходок через слой 4C мы начали выяснять, сохраняются ли такие полоски после закрытия одного глаза и остаются ли нормальными их размеры. Вскоре стало ясно, что слой 4C по-прежнему подразделяется на участки левого и правого глаз, как у нормальных животных, и что клетки в полосках, соединенных с ранее закрытым глазом, остались практически нормальными. Однако последовательности клеток с доминированием закрытого глаза оказались очень короткими, как если бы полоски были аномально узкими (около 0,2 мм вместо 0,4 или 0,5 мм), а полоски, принадлежащие открытому глазу, — соответственно более широкими.
Как только это стало доступным, мы начали использовать методику с инъекцией метки в глаз и транснейронным переносом ее в кору, чтобы получить прямое и наглядное подтверждение сделанных выводов. По прошествии периода депривации длительностью в несколько месяцев мы вводили кошке или обезьяне в тот или в другой глаз радиоактивную аминокислоту. Радиоавтографы показали заметное сужение полосок, связанных с ранее депривированным глазом, и соответственное расширение полосок, принадлежащих нормальному глазу. Микрофотография слева на рис. 145 иллюстрирует результаты инъекции радиоактивной аминокислоты в нормальный глаз. На этом снимке, сделанном, как обычно, в условиях темного поля, представлен срез, параллельный поверхности коры и проходящий через слой 4C. Узкие, прерывающиеся черные полоски соответствуют глазу, который был закрыт, а более широкие светлые (меченые) полоски — открытому глазу, в который ввели метку. На рис. 146 показана обратная картина, полученная после введения метки в ранее закрытый глаз. Этот срез был сделан поперек слоя 4C, поэтому мы видим «полоски» в поперечном сечении.

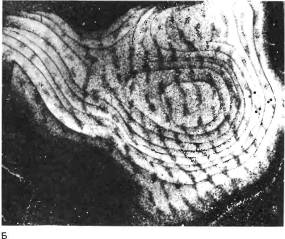
Рис. 145. Срезы зрительной коры макака, правый глаз которого был закрыт с момента рождения до 18-месячного возраста. В левый глаз инъецировали радиоактивную аминокислоту, и неделю спустя приготовили срезы мозга, параллельные поверхности зрительной коры. Кора имеет куполообразную форму, поэтому такие срезы вначале идут тангенциально, а затем образуют кольца (А) все большего диаметра, подобные кольцам луковицы. Эти кольца были вырезаны из микрофотографий и склеены вместе (Б). Впоследствии мы научились расправлять кору перед ее замораживанием, и тогда уже не приходилось разрезать и склеивать фотографии серийных срезов. На обычном радиоавтографе зерна серебра выглядят черными на белом фоне. Мы, однако, применяли метод исследования в темном поле, при котором зерна серебра рассеивают свет и их скопления выглядят как светлые участки. Светлые полоски — места поступления в слой 4C меченых атомов из открытого глаза — расширены, а темные (связанные с закрытым глазом) сильно сужены.

Рис. 146. Другой обезьяне метку ввели в закрытый глаз. Срез сделан в поперечном, а не в тангенциальном направлении. На этом снимке, полученном в темном поле, светлые полоски в слое 4C (которые видны здесь в поперечном сечении) сильно сужены.
Результаты, полученные на слое 4C, подкрепили наши сомнения относительно конкурентной модели, связанные с фактом атрофии клеток коленчатого тела: либо гипотеза о конкуренции была неверна, либо какая-то погрешность была в наших рассуждениях. Оказалось, что погрешность касалась как коленчатого тела, так и коры. В коре нашей ошибкой было предположение, что к моменту, когда мы закрывали глаза новорожденным животным, колонки глазодоминантности были уже вполне сформированы.
Нормальное развитие колонок глазодоминантности
Очевидный способ получения сведений о колонках глазодоминантности у новорожденных животных заключался в проверке распределения волокон, входящих в слой 4C, путем инъекции метки в глаз на первый или второй день жизни. Результат оказался неожиданным. Вместо четких, ясно очерченных полосок в слое 4C мы наблюдали непрерывное распределение метки. Радиоавтограф слева на рис. 147 показывает этот слой в поперечном разрезе, и мы не видим никаких признаков колонок. Только если мы нареза?ли кору параллельно ее поверхности, можно было увидеть слабую волнистость с полумиллиметровыми интервалами, как это показано на радиоавтографе справа. Очевидно, волокна, растущие от коленчатого тела в кору, не разветвляются сразу же к областям, связанным с левым и с правым глазом. Они сначала посылают свои ветви повсюду в радиусе нескольких миллиметров и лишь позднее, приблизительно к моменту рождения животного, втягиваются обратно и окончательно распределяются. Слабая волнистость у новорожденного показывает, что втягивание началось еще до рождения; Пашко Ракич с большими трудностями сумел инъецировать метку в глаз утробного плода обезьяны и выяснил, что оно начинается за несколько недель до рождения. Инъецируя метку в один глаз в различные моменты после рождения, мы смогли легко установить, что в первые две или три недели в слое 4 происходит постепенное втягивание окончаний, так что к четвертой неделе формирование полосок завершается. Узор полосок и их периодичность с шагом 0,8 миллиметра, таким образом, являются врожденными.
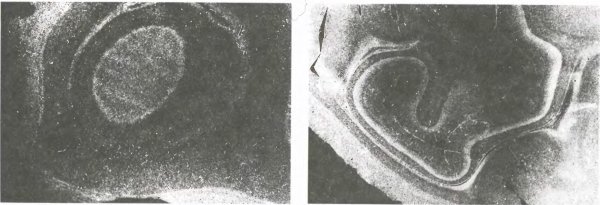
Рис. 147. Слева: поперечный срез слоя 4C коры новорожденного макака после инъекции радиоактивной метки в глаз. Снимок сделан в темном поле, поэтому меченые участки выглядят светлыми. Их непрерывность показывает, что входы от каждого глаза не сгруппированы в полоски, а перемешаны во всем слое. (Белая полоса между слоями 4C, находящимися у поверхности мозга и в глубине, представляет собой белое вещество мозга, заполненное волокнами с меткой, идущими от наружных коленчатых тел.)
Справа: здесь показан срез другого полушария; при изготовлении среза нож слегка коснулся участка стриарной коры, находящегося в глубине. В верхней части слоя 4C можно усмотреть намек на полоски. (Эти полоски находятся в подразделении, связанном с крупноклеточными слоями коленчатого тела.) Более глубинный подслой ? образует непрерывное кольцо вокруг ? и поэтому предположительно позднее дифференцируется по глазодоминантности.
Мы легко подтвердили представление о постнатальном втягивании окончаний, регистрируя ответы слоя 4C обезьян вскоре после рождения. По мере продвижения электрода вдоль этого слоя параллельно его поверхности мы везде смогли вызвать активность от обоих глаз вместо четкого чередования глаз, наблюдаемого у взрослых животных. Карла Шатц продемонстрировала аналогичный процесс развития коленчатого тела у кошки: у утробного плода многие клетки коленчатого тела временно получают входы от обоих глаз, но утрачивают один из входов по мере образования слоистой структуры. Таким образом, и в коре, и в коленчатом теле мы имеем примеры формирования и последующей спонтанной дегенерации синапсов в процессе развития.
Окончательная картина чередования полос для левого и правого глаза в корковом слое 4C развивается нормально даже в том случае, если оба глаза закрыты путем сшивания век; это означает, что надлежащие связи могут создаваться и без участия индивидуального опыта. Мы полагаем, что в процессе развития входные волокна от обоих глаз конкурируют в слое 4C таким образом, что если входы от одного из глаз в каком-то месте преобладают, то преимущество этого глаза имеет тенденцию увеличиваться, а число входов от другого глаза соответственно убывает. Даже небольшое начальное неравенство при этом стремится постепенно возрастать до тех пор, пока всюду в слое 4 в возрасте одного месяца не появятся отчетливые полоски с полным доминированием. При закрытом глазе баланс нарушается, и на границах полосок, где в норме исход борьбы решает прямое столкновение, открытый глаз получает преимущество и одерживает верх, что показано на рис. 148.
Мы не знаем, что? при нормальном развитии приводит к начальному дисбалансу, но при столь неустойчивом равновесии дело может решить даже мельчайшее различие. Почему развивающийся узор должен состоять из параллельных полосок шириной 0,5 мм каждая, остается предметом догадок. Некоторые исследователи предполагают, что аксоны от одного и того же глаза, сближаясь, начинают притягивать друг друга, а от разных глаз — отталкивать друг друга, причем на малых расстояниях отталкивание слабее притяжения, так что притяжение побеждает. С увеличением расстояния притягивающая сила убывает быстрее отталкивающей, так что на некотором удалении побеждает отталкивание. Шириной зоны этих конкурентных взаимодействий и определяются размеры колонок. Согласно математическим расчетам, для получения параллельных полосок (в противовес шахматному порядку или островкам аксонов от левого глаза в сплошной массе аксонов от правого глаза) требуется лишь то, чтобы границы между колонками были как можно короче. Итак, мы имеем возможность объяснить сужение и расширение колонок, показав, что в то время, когда глаз был закрыт в раннем периоде жизни, конкуренция все же была возможной.
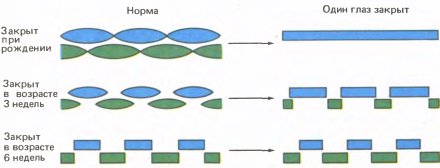
Рис. 148. Эта «конкурентная модель» объясняет разделение волокон четвертого слоя на колонки глазодоминантности. К моменту рождения колонки уже начали формироваться. В норме, если в какой-то точке один глаз хотя бы незначительно доминирует, это оканчивается его полной монополией. Если один глаз при рождении закрыт, то сохранившиеся волокна от открытого глаза в любой данной точке слоя 4 полностью одерживают верх. Волокна от закрытого глаза сохраняются только там, где в момент закрытия у них не было конкурентов.
Тем временем Рэй Гиллери, работавший тогда в Висконсинском университете, предложил правдоподобное объяснение атрофии клеток в коленчатом теле. Изучая наши рисунки, демонстрирующие сморщивание клеток после закрытия одного глаза у кошек, он заметил, что в участках коленчатого тела, наиболее удаленных от средней линии, сморщивание было значительно меньшим; действительно, там — в проекции височной серповидной области — клетки казались вполне нормальными. Здесь отображается боковой край зрительного поля, настолько удаленный от середины, что его может видеть лишь один глаз этой стороны (рис. 149). Мы были огорчены, если не сказать больше, ибо настолько увлеклись измерением клеток с целью подтвердить наши выводы, что просто забыли как следует рассмотреть собственные микрофотографии. Ведь отсутствие атрофии клеток со входами от «височного серпа» означало, что атрофия в других местах коленчатого тела, и в самом деле, могла быть результатом конкуренции со стороны другого глаза.
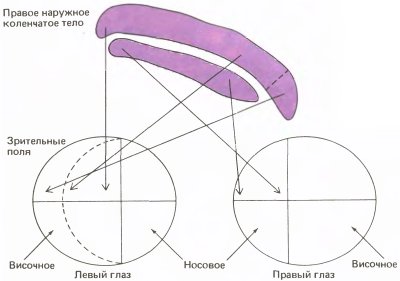
Рис. 149. Различные участки обеих сетчаток проецируются на их собственные зоны в правом коленчатом теле кошки (которое показано здесь в поперечном сечении). Верхний слой коленчатого тела, получающий входные волокна от контралатерального (левого) глаза, нависает над следующим слоем. Свешивающаяся часть получает входы от височного серповидного участка поля зрения, который проецируется на сетчатку только одного глаза. При закрытии этого глаза свешивающаяся часть не атрофируется — предположительно потому, что не встречает конкуренции со стороны другого глаза, в котором соответственного участка нет.
С помощью весьма остроумного эксперимента, показанного на рис. 150, Маррей Шерман и его коллеги представили решающие доказательства роли конкуренции в атрофии клеток коленчатого тела. Сначала они разрушили у котенка крошечный участок одной сетчатки в области бинокулярного поля зрения. Затем они плотно зашили другой глаз. В небольшой зоне того слоя коленчатого тела, на который проецировался глаз с локальным повреждением, была обнаружена сильно выраженная атрофия. Этот результат получили и многие другие исследователи. В слое, имевшем входы от другого, ранее закрытого глаза, как и ожидалось, они тоже наблюдали сморщивание клеток повсюду, кроме зоны, подвергшейся атрофии в слое с проекцией другого глаза. Там, несмотря на отсутствие входных сигналов от глаза, клетки были нормальными. Атрофия, обусловленная закрытием глаза, была предотвращена путем исключения конкуренции. Ясно, что конкуренция не могла происходить в самом коленчатом теле; однако следует помнить, что, хотя тела и дендриты клеток коленчатого тела находились в самом коленчатом теле, большинство окончаний их аксонов были в коре, а там, как я уже говорил, окончания, принадлежащие закрытому глазу, подвергались сильной атрофии. Вывод состоит в том, что при закрытии глаза сморщивание клеток — это результат уменьшения числа поддерживаемых ими аксонных окончаний.
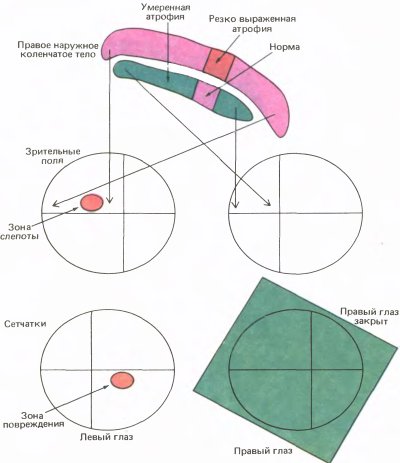
Рис. 150. В 1974 году эксперимент Шермана, Гиллери, Кааса и Сандерсона продемонстрировал значение конкуренции для атрофии клеток наружного коленчатого тела. Если разрушить у котенка небольшой участок левой сетчатки, то в соответствующей части верхнего слоя правого наружного коленчатого тела образуется островок сильно выраженной атрофии. Если затем закрыть правый глаз, то, как и можно было ожидать, слой, расположенный ниже, подвергается атрофии, за исключением участка, лежащего прямо под атрофированным участком верхнего слоя. Этот факт служит сильным доводом в пользу конкурентной природы атрофии, вызываемой закрытием глаза.
Открытие того факта, что при рождении во всем слое 4 без каких-либо перерывов представлены волокна от обоих глаз, оказалось весьма полезным, так как оно объясняло, каким образом могла бы реализоваться конкуренция на синаптическом уровне в структуре, в которой, казалось бы, нет никаких возможностей для взаимодействия глаз. И все-таки проблема может оказаться не такой простой. Если причина изменений в слое 4 — то, что в первые недели после рождения здесь существуют условия для конкуренции, то закрытие глаза в возрасте, когда система еще пластична, а колонки уже разделены, не приводило бы к возникновению изменений. Мы закрывали глаз в возрасте пяти с половиной недель и только через год вводили метку в другой глаз. Результатом было четкое сужение и расширение соответствующих полос. Это как будто бы указывает на то, что помимо дифференцированного втягивания окончаний возможно и прорастание их на новую территорию.
Другие специальные эксперименты с депривацией
Во всех описанных до сих пор исследованиях мы закрывали один или оба глаза или же перереза?ли наружные мышцы одного из глаз. Вскоре во многих лабораториях было проведено множество других экспериментов, включающих чуть ли не все мыслимые виды зрительной депривации. В одном из первых и наиболее интересных экспериментов ставился вопрос, будет ли содержание животного в условиях, позволяющих ему видеть полоски лишь одной ориентации, приводить к утрате клеток, чувствительных ко всем иным ориентациям. В 1970 году Колин Блейкмор и Дж. Ф. Купер из Кембриджского университета ежедневно с раннего возраста показывали котятам в течение нескольких часов чередующиеся черные и белые вертикальные полосы, а в остальное время содержали их в темноте. Результатом было сохранение корковых клеток, реагирующих на вертикальные полосы, и резкое уменьшение числа клеток, предпочитающих другие ориентации. Неясно, перестали ли клетки с исходно промежуточными ориентациями отвечать вовсе или они сменили предпочитаемое направление на вертикальное. В опубликованной в том же году работе Хельмут Хирш и Нико Спинелли использовали очки, позволявшие котенку видеть одним глазом только вертикальные, а другим — только горизонтальные контуры. В результате получилась кора, содержащая клетки с предпочтением вертикалей, клетки с предпочтением горизонталей, но очень мало клеток, предпочитающих наклонные линии. Кроме того, на клетки, активируемые горизонтальными линиями, влиял лишь тот глаз, который подвергался раньше воздействию горизонтальных линий, а на клетки, возбуждаемые вертикальными линиями, — лишь глаз, подвергавшийся воздействию вертикальных линий.
Другие интересные процедуры включали выращивание животных в темном помещении, в котором один или несколько раз в секунду вспыхивал яркий импульс света; он позволял животному увидеть, где оно находится, но должен был сводить к минимуму восприятие любого движения. Результатом этих экспериментов, которые проводили в 1975 году Макс Цинадер, Нэнси Берман и Алан Хейн в Массачусетсском технологическом институте, а также М. Цинадер и Г. Черненко в Далхауси (Галифакс), было уменьшение числа клеток, чувствительных к движению. В другой серии экспериментов, которую начали Ф. Треттер, М. Цинадер и Вольф Зингер в Мюнхене, животным показывали только движение полосок слева направо и получили ожидаемое асимметричное распределение в коре клеток, чувствительных к направлению движения. С большими трудностями и затратами мы с Торстеном Визелом выращивали детеныша обезьяны в комнате, освещаемой лишь длинноволновым красным светом, а затем регистрировали ответы клеток наружных коленчатых тел, чтобы выяснить, не окажется ли здесь необычно мало цветокодирующих нейронов (см. гл. 8). Какой-либо аномалии в коленчатых телах обнаружить не удалось.
Наконец, в последнее десятилетие появилось множество исследований, имеющих целью узнать, содержат ли модифицируемые синапсы такие специальные нейромедиаторы или нейромодуляторы, как норадреналин, а также ацетилхолин или серотонин. Результаты этих исследований представят большой интерес.
Значение результатов депривации в более широкой перспективе
Часто спрашивают, какой цели служит пластичность зрительной системы в ранний период жизни (у человека период чувствительности предположительно составляет четыре — пять лет). У животных, у которых один глаз зашит с момента рождения, территория открытого глаза в слое 4C расширяется; дает ли это какое-либо преимущество открытому глазу? Ответа на этот вопрос еще нет. Трудно представить себе, что острота зрения становится лучше, чем в норме, поскольку нормальная острота, измеряемая офтальмологом с помощью испытательной таблицы, определяется в конечном счете плотностью упаковки рецепторов (см. гл. 3), а она уже лимитирована длиной световых волн.
Как бы то ни было, кажется весьма маловероятным, чтобы пластичность выработалась в ходе эволюции всего лишь на случай возможной утраты глаза детенышем или появления у него косоглазия. Популярная и, несомненно, правдоподобная идея состоит в том, что пластичность обеспечивает тонкую настройку связей, необходимую для восприятия форм, движения и глубины, и что эта настройка осуществляется в основном постнатально под контролем самого зрения. Эта идея привлекательна потому, что способность мозга обучаться может избавить его от необходимости заранее программировать все детали и может обеспечить достаточную гибкость для адаптации к разнообразным условиям среды. Однако для такой идеи пока еще нет убедительных экспериментальных подтверждений. Я лично склонен думать, что в первичной зрительной коре и, быть может, на нескольких последующих уровнях связи полностью определяются генетическими инструкциями. Ясно, что значительная часть этих связей образуется пренатально и, следовательно, без участия сенсорного опыта, и какая бы стратегия ни использовалась для их построения, она могла бы также обеспечивать и их тонкую настройку.
Я не утверждаю, что так же обстоит дело и в других областях коры. Большинство неврологов полагают, что нейронные цепи, ответственные за овладение языком, находятся в основном в коре, — и никто не станет утверждать, что мы рождаемся со знанием тонкостей нашего родного языка. Модифицируемость различных корковых зон и возрастной диапазон, в пределах которого возможны модификации, могут широко варьировать от зоны к зоне, причем первичная зрительная кора с ее минимальной пластичностью, ограниченной наименьшим числом лет постнатального развития, видимо, являет собой крайний случай. На более периферических уровнях зрительной системы, включающих сетчатку, коленчатое тело и первичную кору, пластичность может быть просто побочным продуктом созревания; здесь, я полагаю, определенную роль играет конкуренция, и помехи в ее осуществлении ведут к искажению связей. Но в действительности мы еще не знаем, почему зрительная кора может быть модифицирована в раннем периоде жизни.
Стоит отметить, что все модификации в наших экспериментах явились результатом аномального раннего опыта и привели к образованию аномальных связей. Эксперименты, имевшие целью показать, что обогащенный ранний опыт ведет к усиленному развитию коры или иных структур мозга, не кажутся мне очень убедительными.
Что больше всего поразило нас в экспериментах со зрительной депривацией, так это возможность вызывать заметные физиологические и морфологические изменения в нервной системе без реального физического вмешательства. Давно известно, что клетки в нервной системе могут дегенерировать, если нерв физически перерезать или раздавить, но в описанных мною экспериментах все воздействие сводилось к исключению света, а в опытах с косоглазием вмешательство было еще более тонким. В каждом случае «наказание» более или менее соответствует «преступлению». Исключите форму, и клетки, в норме отвечающие только на формы, перестанут на них отвечать. Измените относительное положение глаз, перерезав одну из мышц, и связи, в норме обслуживающие бинокулярные взаимодействия, будут разорваны. Устраните подвижные стимулы вообще или движение их в определенном направлении, и соответствующие клетки не будут реагировать.
Вряд ли нужно обладать каким-то особым воображением, чтобы подумать о том, что у ребенка, лишенного социальных контактов, или у выращенного в изоляции животного, как в некоторых экспериментах Гарри Харлоу, могут возникнуть аналогичные, столь же реальные изменения в зонах мозга, определяющих эмоции или взаимодействия с другими особями того же вида. Конечно, ни один патолог изменений пока не наблюдал, но ведь и после зрительной депривации в коре тоже нельзя без весьма специальных методов (вроде инъекции метки и глаз) увидеть какие-либо изменения. Когда одни аксоны укорачиваются, а другие удлиняются, вся структура даже при исследовании в электронном микроскопе выглядит точно так же.
Таким образом, возможное значение экспериментов с депривацией выходит далеко за рамки зрительной системы — оно касается также неврологии в целом и в значительной мере психиатрии. Фрейд вполне мог быть прав, связывая психоневрозы с событиями раннего детства, и, учитывая его неврологическое образование, я думаю, он был бы в восторге от мысли, что подобные события могут приводить к заметным гистологическим или гистохимическим изменениям в реальном физическом мозгу.
Глаз, мозг, зрение
Дэвид Хантер Хьюбел; David Hunter Hubel;
Дек 30 2017